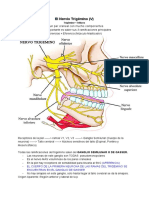
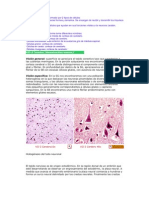
You might also like
- Manual Práctico de AstrologíaDocument228 pagesManual Práctico de AstrologíaIvan Larreynaga82% (11)
- Pared de Gram NegativosDocument31 pagesPared de Gram NegativosDarla SanchezNo ratings yet
- Complejos de UniónDocument7 pagesComplejos de UniónAnalia Moraes100% (1)
- Plan Anual CCNN Noveno EgbDocument38 pagesPlan Anual CCNN Noveno EgbCarlos Oswaldo García Paredes100% (2)
- Glosario de EmbriologiaDocument12 pagesGlosario de EmbriologiaMayer TerronesNo ratings yet
- Agosto - 4to Grado Ciencias Naturales (2018-2019)Document4 pagesAgosto - 4to Grado Ciencias Naturales (2018-2019)Fernanda Loredl'a100% (1)
- Embriologia Dentaria OdontogenesisDocument12 pagesEmbriologia Dentaria OdontogenesisDarío Neira100% (1)
- Botánica Agrícola, Raíz 2014Document33 pagesBotánica Agrícola, Raíz 2014Liz Bonifacio Durant100% (1)
- Mecanismos de Evasion de La Respuesta InmuneDocument8 pagesMecanismos de Evasion de La Respuesta InmuneNnznznznNo ratings yet
- TriangulosAnatomiaDocument3 pagesTriangulosAnatomiaJairo Kevin Bautista Castillo100% (2)
- Técnicas de muestreo y monitoreo de floraDocument26 pagesTécnicas de muestreo y monitoreo de floraLuisMiguelAlva67% (6)
- Practica de Embriologia DentariaDocument12 pagesPractica de Embriologia DentariaGerardo Bardales ArgüellesNo ratings yet
- Gerencia y productos de explosivos para mineríaDocument33 pagesGerencia y productos de explosivos para mineríaelias100% (1)
- Propiedades Físicas y Mecánicas de Los Materiales Dentales y Su Relación Con Los Tejidos Dentarios. - CompressedDocument15 pagesPropiedades Físicas y Mecánicas de Los Materiales Dentales y Su Relación Con Los Tejidos Dentarios. - CompressedDayana AyalaNo ratings yet
- Relaciones de La Célula Con Su EntornoDocument4 pagesRelaciones de La Célula Con Su EntornoAndrea Mendoza0% (1)
- Odonto GenesisDocument5 pagesOdonto GenesisFiamma AguirreNo ratings yet
- Anatomía Canino 40Document2 pagesAnatomía Canino 40Nancy OrtegaNo ratings yet
- Cartílago HialinoDocument3 pagesCartílago HialinoSofia DuranNo ratings yet
- Tejido Conectivo LaxoDocument2 pagesTejido Conectivo LaxoRodrigo PerezNo ratings yet
- CATM Desarrollo PrenatalDocument7 pagesCATM Desarrollo PrenatalKarina Marly SantanaNo ratings yet
- DGB y DGT PresentaciónDocument36 pagesDGB y DGT PresentaciónEdgaris Sc'r0% (1)
- 6to Informe - Lab BiologiaDocument9 pages6to Informe - Lab Biologiapaola86% (14)
- Complejo Dentino - PulparDocument26 pagesComplejo Dentino - PulparLucia ManzanoNo ratings yet
- PERIO Saco PeriodontalDocument6 pagesPERIO Saco PeriodontalKiara Bello100% (1)
- Ligamento periodontal: estructura y funcionesDocument50 pagesLigamento periodontal: estructura y funcionesGabriel GorotizaNo ratings yet
- Glosario de OrganogénesisDocument6 pagesGlosario de OrganogénesisMario José QuesadaNo ratings yet
- Par Craneal 5 PDFDocument8 pagesPar Craneal 5 PDFAlicia Trimmiño JaramilloNo ratings yet
- Preguntas Glndulas SalivalesDocument9 pagesPreguntas Glndulas SalivalesSebastián Estay SaavedraNo ratings yet
- Arteria Maxilar InternaDocument4 pagesArteria Maxilar InternaPablo DBNo ratings yet
- Origen y trayecto de las arterias y venas de la cabeza y el cuelloDocument45 pagesOrigen y trayecto de las arterias y venas de la cabeza y el cuelloArturo Oscar Martinez Aguilar100% (3)
- Conclusion Tejido EpitelialDocument3 pagesConclusion Tejido EpitelialMaroly Alcivar100% (1)
- Histogénesis Del EsmalteDocument3 pagesHistogénesis Del EsmalteHamilton AndradeNo ratings yet
- Histologia EsmalteDocument27 pagesHistologia EsmalteAngie Sofia Sanchez TatisNo ratings yet
- Ejercicios de OxidacionDocument3 pagesEjercicios de OxidacionAngel ManriqueNo ratings yet
- Embriogenesis de La Cuarta Semana A La Octava SemanaDocument55 pagesEmbriogenesis de La Cuarta Semana A La Octava SemanaZOfii SnhzNo ratings yet
- Amelogenesis GRAN MARISCAL DE AYACUCHODocument4 pagesAmelogenesis GRAN MARISCAL DE AYACUCHODoomVoiceNo ratings yet
- Complejo dentino-pulpar: estructura y funciónDocument39 pagesComplejo dentino-pulpar: estructura y funciónJuan BernalNo ratings yet
- Estructura, función e histología del tejido linfoide asociado a mucosa (MALTDocument2 pagesEstructura, función e histología del tejido linfoide asociado a mucosa (MALTYuki Chari Moco Saray50% (4)
- PAUSAS ACTIVAS - Ergonomia DentalDocument7 pagesPAUSAS ACTIVAS - Ergonomia DentalEnrique GamarraNo ratings yet
- Guia 05 Tejido Conectivo PDFDocument8 pagesGuia 05 Tejido Conectivo PDFJuan Carlos Robayo100% (4)
- Practica 4Document7 pagesPractica 4Amisadai Cruz ReyesNo ratings yet
- Biotipo ConstitucionalDocument6 pagesBiotipo ConstitucionalcarlodNo ratings yet
- Homeostasis Del Sodio y AguaDocument25 pagesHomeostasis Del Sodio y AguaFdo RM100% (1)
- Musculos InfrahioideosDocument60 pagesMusculos Infrahioideosloquito2012100% (1)
- Selección Dientes ProteicosDocument15 pagesSelección Dientes ProteicosartvroandradeNo ratings yet
- Tema 6.2. Arteria Maxilar InternaDocument20 pagesTema 6.2. Arteria Maxilar InternaFrank Santos HerediaNo ratings yet
- El complejo dentinopulpar: estructura, funciones y característicasDocument5 pagesEl complejo dentinopulpar: estructura, funciones y característicasAlexander Ruiz100% (1)
- Formación de La Dentina CircumpulparDocument3 pagesFormación de La Dentina CircumpulparMaria RojasNo ratings yet
- Biofisica 2Document15 pagesBiofisica 2AarónNo ratings yet
- Funcion Del Colesterol y La ProteinasDocument3 pagesFuncion Del Colesterol y La ProteinasJoseCarlosNo ratings yet
- Articulacion Alveolo DentariaDocument8 pagesArticulacion Alveolo DentariaNegrita Cueva100% (1)
- La SalivaDocument6 pagesLa SalivaBety Jim AraNo ratings yet
- Para Tiro IdeaDocument7 pagesPara Tiro IdeaJochiJaramilloNo ratings yet
- Disco BilaminarDocument43 pagesDisco BilaminarArtuRock182No ratings yet
- Tema 1Document23 pagesTema 1MaPi OhhNo ratings yet
- Transporte Mediado Por ProteínasDocument8 pagesTransporte Mediado Por ProteínasMaite Celi LoayzaNo ratings yet
- Hueso Esfenoides Informe.Document7 pagesHueso Esfenoides Informe.Dania Baculima100% (1)
- Membrana Plasmatica (Informe)Document9 pagesMembrana Plasmatica (Informe)Lucio PerezNo ratings yet
- 07.FILAMENTOS INTERMEDIOS - CooperDocument8 pages07.FILAMENTOS INTERMEDIOS - CooperNatalia DonatoNo ratings yet
- Tipos de uniones celulares: características y funcionesDocument14 pagesTipos de uniones celulares: características y funcionesGabriela EscalanteNo ratings yet
- Gartner, Leslie P. (19 Sistema Urinario)Document24 pagesGartner, Leslie P. (19 Sistema Urinario)Ardila NubiaNo ratings yet
- Examen EmbriologiaDocument4 pagesExamen EmbriologiaMartha RLNo ratings yet
- Tejido nervioso: neuronas y neurogliasDocument2 pagesTejido nervioso: neuronas y neurogliasjhackisNo ratings yet
- PDF Las Interacciones de Las Celulas Entre Si Grupo 5 DDDocument7 pagesPDF Las Interacciones de Las Celulas Entre Si Grupo 5 DDpilar morochoNo ratings yet
- Guia Adhesion Celular 2011Document4 pagesGuia Adhesion Celular 2011Clarys DiazNo ratings yet
- GUIA DE LAS ESTRUCTURAS DE ADHESIÓN CELULARDocument4 pagesGUIA DE LAS ESTRUCTURAS DE ADHESIÓN CELULARCarolina Severino0% (1)
- Teoría Piaget y VygotskyDocument2 pagesTeoría Piaget y VygotskyAriel PincayNo ratings yet
- Polímeros: Importancia de los monómerosDocument29 pagesPolímeros: Importancia de los monómerosAriel PincayNo ratings yet
- Polímeros: Importancia de los monómerosDocument29 pagesPolímeros: Importancia de los monómerosAriel PincayNo ratings yet
- Polímeros: Importancia de los monómerosDocument29 pagesPolímeros: Importancia de los monómerosAriel PincayNo ratings yet
- Análisis en Los Entornos de La EcologíaDocument4 pagesAnálisis en Los Entornos de La EcologíaAriel PincayNo ratings yet
- Evaluación InstitucionalDocument4 pagesEvaluación InstitucionalAriel PincayNo ratings yet
- Análisis en Los Entornos de La EcologíaDocument4 pagesAnálisis en Los Entornos de La EcologíaAriel PincayNo ratings yet
- Reactivo Evaluacion FinalDocument8 pagesReactivo Evaluacion FinalAriel PincayNo ratings yet
- Normas de Seguridad Generales para LaboratorioDocument4 pagesNormas de Seguridad Generales para LaboratorioAriel PincayNo ratings yet
- Evaluacion Formativa CasoDocument3 pagesEvaluacion Formativa CasoAriel PincayNo ratings yet
- Correcciones de La Inclinación y Declinación MagnéticaDocument10 pagesCorrecciones de La Inclinación y Declinación MagnéticaLuisSaavedraFloresNo ratings yet
- Viga T InvertidaDocument68 pagesViga T InvertidaGonzalo VegaNo ratings yet
- El Mendigo de AlmasDocument8 pagesEl Mendigo de AlmasErick LSNo ratings yet
- Las Dos Caras de La CienciaDocument2 pagesLas Dos Caras de La CienciaSergio RozoNo ratings yet
- 100 Preguntas de Geologia y MineralogiaDocument2 pages100 Preguntas de Geologia y MineralogiaSantiago Parra JimenezNo ratings yet
- MEMORIA DESCRIPTIVA OBRAS DE ARTE Km. 35+000 - 153+500Document7 pagesMEMORIA DESCRIPTIVA OBRAS DE ARTE Km. 35+000 - 153+500sadhafzNo ratings yet
- Guía 35 EL-31 EGRESADOS Estrategias para Int Textos Que Plantean Distintos Mundos Literarios - PRO PDFDocument16 pagesGuía 35 EL-31 EGRESADOS Estrategias para Int Textos Que Plantean Distintos Mundos Literarios - PRO PDFClaudia CáceresNo ratings yet
- Extracción ADN fresasDocument3 pagesExtracción ADN fresasSaidyVillalba100% (1)
- Ingredientes y Aditivos Usados en La Industria Carnica (Autoguardado)Document23 pagesIngredientes y Aditivos Usados en La Industria Carnica (Autoguardado)YUNIORMONZONNo ratings yet
- Problemas Repaso DinamicaDocument14 pagesProblemas Repaso DinamicaDiego AzuaraNo ratings yet
- La Maldicion de La Sangre 01 - El Abogad - Gherbod FlemingDocument210 pagesLa Maldicion de La Sangre 01 - El Abogad - Gherbod Fleminglasartenporelmango100% (1)
- Perspectivas Cognoscitivas Del AprendizajeDocument14 pagesPerspectivas Cognoscitivas Del AprendizajeSandra Herrera Gonzalez100% (3)
- Texto Arg. Entrega 1Document7 pagesTexto Arg. Entrega 1PAULA ANDREA VANEGAS SILVANo ratings yet
- Guia de Obra Dramatica PDFDocument6 pagesGuia de Obra Dramatica PDFJenny Gisela RuzNo ratings yet
- Fuerzas concurrentes laboratorio físicaDocument32 pagesFuerzas concurrentes laboratorio físicaJhonTrujilloNo ratings yet
- Proyecto de Investigacion TuristicaDocument7 pagesProyecto de Investigacion TuristicaEliana OrtegaNo ratings yet
- Biografias de Matematicos de Algebra de BaldorDocument11 pagesBiografias de Matematicos de Algebra de BaldorAlexis100% (1)
- M 3Document3 pagesM 3Francisco Sanchez RasgadoNo ratings yet
- 0170 Titulo XV. Normas de Seguridad Aplicables A Faenas Mineras Que Indica 594-635Document14 pages0170 Titulo XV. Normas de Seguridad Aplicables A Faenas Mineras Que Indica 594-635Kenny CasillaNo ratings yet
- UNIDADESDocument13 pagesUNIDADESAyrtonNo ratings yet
- Solucionario Semana05 Ord2012 IDocument99 pagesSolucionario Semana05 Ord2012 Iobdzcopk2No ratings yet
- Levantamiento Pilas 19 01 17Document1 pageLevantamiento Pilas 19 01 17MiguelAngelAceitonVegaNo ratings yet
- Practica 3 Bioquimica FinalDocument12 pagesPractica 3 Bioquimica FinalAlex CustomNo ratings yet
- Semana 01 EdafologiaDocument34 pagesSemana 01 EdafologiaAnibal PariaNo ratings yet