You might also like
- Ang Pasko Ay SumapitDocument2 pagesAng Pasko Ay SumapitJhoana Ofrasio90% (10)
- Now We Remain PDFDocument4 pagesNow We Remain PDFJomari M. Marasigan100% (1)
- Kathang Isip: Ben & BenDocument6 pagesKathang Isip: Ben & BenJomari M. Marasigan100% (2)
- Asme NM.2-2018Document126 pagesAsme NM.2-2018aneeshjokay75% (4)
- Baby Its Cold Outside PDFDocument4 pagesBaby Its Cold Outside PDFJomari M. MarasiganNo ratings yet
- Can T Help Falling in Love PDFDocument3 pagesCan T Help Falling in Love PDFJomari M. MarasiganNo ratings yet
- 4-Week Weight Loss ChallengeDocument6 pages4-Week Weight Loss ChallengeTammy JacksonNo ratings yet
- S6MT 1Q w1 3 MELC1 SLM MIXTURES FinalCopy09082020Document26 pagesS6MT 1Q w1 3 MELC1 SLM MIXTURES FinalCopy09082020Rona Dindang100% (1)
- Age Discrimination PDFDocument20 pagesAge Discrimination PDFMd. Rezoan ShoranNo ratings yet
- Objection HandlingDocument3 pagesObjection HandlingNabin GaraiNo ratings yet
- 01 Slug CatchersDocument23 pages01 Slug CatchersMohamed Sahnoun100% (2)
- PE 560 Atomic Absorption Spectrophotometer ManualDocument176 pagesPE 560 Atomic Absorption Spectrophotometer ManualDavid Allan100% (1)
- Holy Cross Academy Quarterly Cookery ExamDocument4 pagesHoly Cross Academy Quarterly Cookery ExamAlle Eiram Padillo95% (21)
- Brazilian Journal of MicrobiologyDocument11 pagesBrazilian Journal of MicrobiologySukoNo ratings yet
- S2589014X21001419Document7 pagesS2589014X21001419Jefferson ContrerasNo ratings yet
- Spirulina Growth Using Fresh Water Supplemented with NutrientsDocument6 pagesSpirulina Growth Using Fresh Water Supplemented with NutrientsStefano CarreñoNo ratings yet
- Effects of Nitrate Toxicity in The Pacific White Shrimp, Litopenaeus Vannamei, Reared With Biofloc Technology (BFT)Document13 pagesEffects of Nitrate Toxicity in The Pacific White Shrimp, Litopenaeus Vannamei, Reared With Biofloc Technology (BFT)MillerCasTroNo ratings yet
- 1 s2.0 S096085241931291X MainDocument8 pages1 s2.0 S096085241931291X MainShabrina Nurul FatimahNo ratings yet
- Spirulina PhotobioreactorDocument7 pagesSpirulina PhotobioreactorManuel Cortés LozanoNo ratings yet
- Soletto 2005Document8 pagesSoletto 2005Jefferson ContrerasNo ratings yet
- Chapter3 PDFDocument32 pagesChapter3 PDFmadhujayarajNo ratings yet
- Bilbao, 2016. Haematococcus Pluvialisas A Source of Fatty Acids and Phytosterols Potential Nutritional and Biological ImplicationsDocument12 pagesBilbao, 2016. Haematococcus Pluvialisas A Source of Fatty Acids and Phytosterols Potential Nutritional and Biological ImplicationsM Rizki Ariansyah putraNo ratings yet
- Different Nitrogen Sources and Growth Responses ofDocument5 pagesDifferent Nitrogen Sources and Growth Responses ofJuan David LsNo ratings yet
- Astaxanthin Production From A New Strain ofDocument9 pagesAstaxanthin Production From A New Strain ofamensetNo ratings yet
- Molecular Characterization of The Nitrifying Bacterial ConsortiaDocument8 pagesMolecular Characterization of The Nitrifying Bacterial ConsortiaSamuel Pereira SanandresNo ratings yet
- A-heavy-metal-tolerant-novel-bacterium,-Bacillus-malikii-sp.-nov.,-isolated-from-tannery-effluent-wastewater_2015_Antonie-van-Leeuwenhoek,-International-Journal-of-General-and-Molecular-Microbiology.pdfDocument12 pagesA-heavy-metal-tolerant-novel-bacterium,-Bacillus-malikii-sp.-nov.,-isolated-from-tannery-effluent-wastewater_2015_Antonie-van-Leeuwenhoek,-International-Journal-of-General-and-Molecular-Microbiology.pdfCristian SalgadoNo ratings yet
- Received: Accepted:: Afegbua, S.L., Jones, A.L., Sani, Y.M., Okoro, C. and Goodfellow, MDocument9 pagesReceived: Accepted:: Afegbua, S.L., Jones, A.L., Sani, Y.M., Okoro, C. and Goodfellow, MUMYU Journal of Microbiology Research (UJMR)No ratings yet
- Pengaruh Konsentrasi Natrium Nitrat Dan Natrium Dehidrogen Fosfat Pada Media Walne Terhadap Konsentrasi Biomassa Dan Protein Nannochloropsis OculataDocument10 pagesPengaruh Konsentrasi Natrium Nitrat Dan Natrium Dehidrogen Fosfat Pada Media Walne Terhadap Konsentrasi Biomassa Dan Protein Nannochloropsis OculatadikimstNo ratings yet
- Model-2 PHB Azohydromonas LataDocument11 pagesModel-2 PHB Azohydromonas LatamtlopezNo ratings yet
- International Biodeterioration & BiodegradationDocument8 pagesInternational Biodeterioration & BiodegradationDuke Renovar MonsaleNo ratings yet
- Effects of Salinity On Growth and Lipid Accumulation of Biofuel Microalga Nannochloropsis Salina and Invading Organisms - Elsevier Enhanced ReaderDocument10 pagesEffects of Salinity On Growth and Lipid Accumulation of Biofuel Microalga Nannochloropsis Salina and Invading Organisms - Elsevier Enhanced ReaderBrigita de BrillarNo ratings yet
- Sampaio Et Al 2007Document9 pagesSampaio Et Al 2007Bruna FacundesNo ratings yet
- Optimization of Biomass, Total Carotenoids and Astaxanthin Production in Haematococcus Pluvialis Flotow Strain Steptoe (Nevada, USA) Under Laboratory ConditionsDocument15 pagesOptimization of Biomass, Total Carotenoids and Astaxanthin Production in Haematococcus Pluvialis Flotow Strain Steptoe (Nevada, USA) Under Laboratory ConditionsNelson Gonzalez MedelNo ratings yet
- 12 - Microalgae Population Dynamics in PhotobioreactorsDocument10 pages12 - Microalgae Population Dynamics in PhotobioreactorsAnthony Emilio Roman VegaNo ratings yet
- (2006) - Commercial Applications of MicroalgaeDocument39 pages(2006) - Commercial Applications of MicroalgaeamensetNo ratings yet
- 2022 Faisal Plant Physiology and BiochemistryDocument10 pages2022 Faisal Plant Physiology and Biochemistryrashid.ssNo ratings yet
- Effects of Carbon Source On The Formation, Stability, Bioactivity andDocument10 pagesEffects of Carbon Source On The Formation, Stability, Bioactivity andsamuel campos carvalhoNo ratings yet
- Mera 2016Document14 pagesMera 2016NadineNo ratings yet
- Dovis 2014Document8 pagesDovis 2014raíssa ,No ratings yet
- Validation of A Spectrophotometric Methodology For A Rapid Iodine AnalysisDocument8 pagesValidation of A Spectrophotometric Methodology For A Rapid Iodine AnalysisDuy Thịnh Trần NguyễnNo ratings yet
- Research Proposal/Synopsis For M.S Thesis Department of BiologyDocument22 pagesResearch Proposal/Synopsis For M.S Thesis Department of BiologyBasit KhanNo ratings yet
- Bioresource TechnologyDocument8 pagesBioresource TechnologyDaniela BravoNo ratings yet
- Biofixation of Carbon Dioxide by Spirulina Sp. and Scenedesmus Obliquus Cultivated in A Three-Stage Serial Tubular Photobioreactor PDFDocument7 pagesBiofixation of Carbon Dioxide by Spirulina Sp. and Scenedesmus Obliquus Cultivated in A Three-Stage Serial Tubular Photobioreactor PDFAna Maria AcevedoNo ratings yet
- Tekaya2017 Article ArbuscularMycorrhizalFungusRhiDocument9 pagesTekaya2017 Article ArbuscularMycorrhizalFungusRhiImed CheraiefNo ratings yet
- ZarroukksDocument9 pagesZarroukksKartikkeyyan LoganathanNo ratings yet
- CRF Sorbate and Benzoate ReviewDocument14 pagesCRF Sorbate and Benzoate Reviewaryamanmadan10No ratings yet
- Autotrophic Growth and Carotenoid Production of HP in A 30 Liter Air Lift PhotobioreactorDocument6 pagesAutotrophic Growth and Carotenoid Production of HP in A 30 Liter Air Lift PhotobioreactorcikpayungNo ratings yet
- SOD Gel Stress LycopersiconDocument14 pagesSOD Gel Stress LycopersiconeruditeramanaNo ratings yet
- Biocatalysis and Agricultural BiotechnologyDocument7 pagesBiocatalysis and Agricultural BiotechnologyLydiaNo ratings yet
- Protein Extraction From Algae MethodDocument14 pagesProtein Extraction From Algae MethodImran NajeebNo ratings yet
- Acta TropicaDocument6 pagesActa TropicaSiva PrasadNo ratings yet
- Stable Isotopes Indicate Individual Level Trophic Diversity in Freshwater GastropodDocument5 pagesStable Isotopes Indicate Individual Level Trophic Diversity in Freshwater GastropodMarco Antonio SolisNo ratings yet
- Effects of PH On Nitrogen Transformations in Media-Based Aquaponics (Bioresource Technology) (2016)Document7 pagesEffects of PH On Nitrogen Transformations in Media-Based Aquaponics (Bioresource Technology) (2016)Ash RafiNo ratings yet
- 1 s2.0 S0981942816301917 MainDocument11 pages1 s2.0 S0981942816301917 MainBlue SkyNo ratings yet
- Sinha, Parli - 2020 - Siderophore Production by Bacteria Isolated From Mangrove Sediments A Microcosm Study PDFDocument9 pagesSinha, Parli - 2020 - Siderophore Production by Bacteria Isolated From Mangrove Sediments A Microcosm Study PDFalokNo ratings yet
- 17 Full PDFDocument9 pages17 Full PDFPriscilia YuniarNo ratings yet
- 2011 Guillncasla PCA E-BeamDocument9 pages2011 Guillncasla PCA E-BeamAn Bùi ĐứcNo ratings yet
- Bacillus Subtil Is I JB Pas 20174239Document12 pagesBacillus Subtil Is I JB Pas 20174239Zahra Nur AzizaNo ratings yet
- Diversity of Bacterial Strains Degrading Hexadecane in Relation To The Mode of Substrate UptakeDocument8 pagesDiversity of Bacterial Strains Degrading Hexadecane in Relation To The Mode of Substrate Uptakesaiful2016No ratings yet
- Semicontinuous Cultivation of Spirulina platensis in a Closed PhotobioreactorDocument7 pagesSemicontinuous Cultivation of Spirulina platensis in a Closed PhotobioreactorR_M_M_No ratings yet
- KNO3makale PDFDocument9 pagesKNO3makale PDFTrishia PentecostesNo ratings yet
- A Low-Tech Bioreactor System For The Enrichment and Production of Ureolytic MicrobesDocument8 pagesA Low-Tech Bioreactor System For The Enrichment and Production of Ureolytic MicrobesAfshaNo ratings yet
- Ajol File Journals - 90 - Articles - 135160 - Submission - Proof - 135160 1069 362513 1 10 20160509Document7 pagesAjol File Journals - 90 - Articles - 135160 - Submission - Proof - 135160 1069 362513 1 10 20160509busra.arslanNo ratings yet
- Application of The Synechococcus NirA Promoter ToDocument8 pagesApplication of The Synechococcus NirA Promoter ToEduardo MoreiraNo ratings yet
- Aquaculture and Fisheries: Guozhi Luo, Wenqing Li, Hongxin Tan, Xiaoqing ChenDocument7 pagesAquaculture and Fisheries: Guozhi Luo, Wenqing Li, Hongxin Tan, Xiaoqing ChenBD MUG HOUSE WOODNo ratings yet
- Characterizations of Purple Non-Sulfur Bacteria Isolated From Paddy FieldsDocument10 pagesCharacterizations of Purple Non-Sulfur Bacteria Isolated From Paddy Fieldsdaniela.aparecidaNo ratings yet
- Genotoxicity of food preservatives sodium and potassium benzoateDocument7 pagesGenotoxicity of food preservatives sodium and potassium benzoateYono FabiandiNo ratings yet
- tmp4D6B TMPDocument9 pagestmp4D6B TMPFrontiersNo ratings yet
- j.foodchem.2011.06.023Document8 pagesj.foodchem.2011.06.023silviofratariNo ratings yet
- Algal Sulfur Compound Release in Response to Light RegimesDocument8 pagesAlgal Sulfur Compound Release in Response to Light RegimesErika M. Sánchez MaquiNo ratings yet
- Spirulina Maxima: Production of Phenolic Compounds From MicroalgaeDocument9 pagesSpirulina Maxima: Production of Phenolic Compounds From MicroalgaeMaita NouhaNo ratings yet
- Molecular Breeding and Genetics of Applied MicroorganismsFrom EverandMolecular Breeding and Genetics of Applied MicroorganismsKenji SakaguchiNo ratings yet
- OfficialdismpretenderDocument8 pagesOfficialdismpretenderJomari M. MarasiganNo ratings yet
- Fancy - Twice1 PDFDocument5 pagesFancy - Twice1 PDFJomari M. MarasiganNo ratings yet
- The Social and Economic Impact of Biofortification Through Genetic ModificationDocument8 pagesThe Social and Economic Impact of Biofortification Through Genetic ModificationJomari M. MarasiganNo ratings yet
- Eds 140 Final Paper GuidelinesDocument2 pagesEds 140 Final Paper GuidelinesJomari M. MarasiganNo ratings yet
- Fancy - Twice PDFDocument5 pagesFancy - Twice PDFJomari M. MarasiganNo ratings yet
- Fancy - TwiceDocument5 pagesFancy - TwiceJomari M. MarasiganNo ratings yet
- 3058806-Christmas Song PDFDocument2 pages3058806-Christmas Song PDFJomari M. MarasiganNo ratings yet
- Banana 2nd Article RahulDocument6 pagesBanana 2nd Article RahulThai Minh TranNo ratings yet
- Direct PCRDocument3 pagesDirect PCRJomari M. MarasiganNo ratings yet
- Screening Musa Genotypes For Banana Bunchy Top Disease Resistance in BurundiDocument11 pagesScreening Musa Genotypes For Banana Bunchy Top Disease Resistance in BurundiJomari M. MarasiganNo ratings yet
- A Dozen A Day Book 1 PDFDocument29 pagesA Dozen A Day Book 1 PDFJomari M. Marasigan80% (5)
- Mommy PrintDocument6 pagesMommy PrintJomari M. MarasiganNo ratings yet
- Canon in D Major PDFDocument2 pagesCanon in D Major PDFJomari M. MarasiganNo ratings yet
- Alkali TRMT For PCRDocument2 pagesAlkali TRMT For PCRJomari M. MarasiganNo ratings yet
- Acta Horticultura BBTVMolinaetal 2009Document7 pagesActa Horticultura BBTVMolinaetal 2009Jomari M. MarasiganNo ratings yet
- 010 FilomenaSta - Cruzetal.article Vol.99No.1March20161Document12 pages010 FilomenaSta - Cruzetal.article Vol.99No.1March20161Jomari M. MarasiganNo ratings yet
- Fly Me To The MoonDocument1 pageFly Me To The MoonJomari M. MarasiganNo ratings yet
- 010 FilomenaSta - Cruzetal.article Vol.99No.1March20161Document12 pages010 FilomenaSta - Cruzetal.article Vol.99No.1March20161Jomari M. MarasiganNo ratings yet
- Bert Homie Up MBDocument4 pagesBert Homie Up MBJomari M. MarasiganNo ratings yet
- Sample Scientific Paper 01Document14 pagesSample Scientific Paper 01Jomari M. MarasiganNo ratings yet
- Banana Bunchy Top Virus and The Bunchy Top Disease: Journal of General Plant Pathology December 2015Document13 pagesBanana Bunchy Top Virus and The Bunchy Top Disease: Journal of General Plant Pathology December 2015Jomari M. MarasiganNo ratings yet
- Testblueprint Nasc3 1exam OnlyDocument1 pageTestblueprint Nasc3 1exam OnlyJomari M. MarasiganNo ratings yet
- I Can't Help Falling in Love: Wise Men Say, Ly OnDocument4 pagesI Can't Help Falling in Love: Wise Men Say, Ly OnJomari M. MarasiganNo ratings yet
- 1000010684072Document7 pages1000010684072Jomari M. MarasiganNo ratings yet
- TSB 20230831 - Reconnect FATL WM Error Code & Troubleshooting GuideDocument6 pagesTSB 20230831 - Reconnect FATL WM Error Code & Troubleshooting GuideNavjot KaurNo ratings yet
- IC 33 Question PaperDocument12 pagesIC 33 Question PaperSushil MehraNo ratings yet
- Almond Milk Case Study Executive SummaryDocument19 pagesAlmond Milk Case Study Executive Summarygauthamsindia307No ratings yet
- Flashover Influence of Fog Rate On The Characteristics of Polluted Silicone Rubber InsulatorsDocument5 pagesFlashover Influence of Fog Rate On The Characteristics of Polluted Silicone Rubber InsulatorsdaaanuNo ratings yet
- Yanagiba Sharpening: Everything You Need To KnowDocument16 pagesYanagiba Sharpening: Everything You Need To KnowT ChenNo ratings yet
- 2005-05-12Document18 pages2005-05-12The University Daily KansanNo ratings yet
- AnxietyDocument5 pagesAnxietydrmadankumarbnysNo ratings yet
- Abdul Khaliq - Good Governance (GG)Document15 pagesAbdul Khaliq - Good Governance (GG)Toorialai AminNo ratings yet
- ĐỀ THI THU TNTHPT SỐ 17Document4 pagesĐỀ THI THU TNTHPT SỐ 17Nguyên Hà NguyễnNo ratings yet
- Introduction To Integrative Homeopathy - Bob LeckridgeDocument16 pagesIntroduction To Integrative Homeopathy - Bob LeckridgeBob LeckridgeNo ratings yet
- Health Programs Activities Timeframe Expected Output Child CareDocument3 pagesHealth Programs Activities Timeframe Expected Output Child CareC SamNo ratings yet
- Sampling & Data AssayDocument22 pagesSampling & Data AssayFerdinand SiahaanNo ratings yet
- Secondary AssessmentsDocument12 pagesSecondary Assessmentsapi-338389967No ratings yet
- Lesson 8 - Philippine Disaster Risk Reduction and Management SystemDocument11 pagesLesson 8 - Philippine Disaster Risk Reduction and Management SystemMary Joy CuetoNo ratings yet
- Seguridad Boltec Cable PDFDocument36 pagesSeguridad Boltec Cable PDFCesar QuintanillaNo ratings yet
- EDSP Quantitative and Qualitative FormDocument2 pagesEDSP Quantitative and Qualitative FormTalal SultanNo ratings yet
- Cd6352 Lee Swee LeongDocument24 pagesCd6352 Lee Swee LeongFrea Marie SarabiaNo ratings yet
- Calculation of Lightning and Switching Overvoltages Transferred Through Power TransformerDocument9 pagesCalculation of Lightning and Switching Overvoltages Transferred Through Power TransformerBožidar Filipović-GrčićNo ratings yet
- Final TLE9 Nail Care9 Q1 Module 3Document20 pagesFinal TLE9 Nail Care9 Q1 Module 3Ma. Andrea LagmanNo ratings yet
- The Congressional Committee and Philippine Policymaking: The Case of The Anti-Rape Law - Myrna LavidesDocument29 pagesThe Congressional Committee and Philippine Policymaking: The Case of The Anti-Rape Law - Myrna LavidesmarielkuaNo ratings yet
- Causes of Failure of Earth DamsDocument11 pagesCauses of Failure of Earth DamsThéoneste NSANZIMFURANo ratings yet
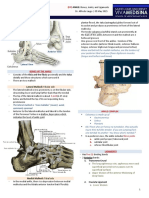
- (9F) Ankle - Bones, Joints, Tendons and LigamentsDocument4 pages(9F) Ankle - Bones, Joints, Tendons and LigamentsJeffrey RamosNo ratings yet