You might also like
- Enzimas OdontologiaDocument52 pagesEnzimas Odontologiakaren ancasiNo ratings yet
- Músculos Masticadores. Músculos Que Mueven La Mandíbula y Fijan El Hioides. Músculos CutáneosDocument52 pagesMúsculos Masticadores. Músculos Que Mueven La Mandíbula y Fijan El Hioides. Músculos CutáneosNoelia JavielNo ratings yet
- Endodoncia Vs PeriodonciaDocument10 pagesEndodoncia Vs PeriodonciacarolinaNo ratings yet
- Quistes Odontogenicos MexicoDocument8 pagesQuistes Odontogenicos MexicoDanae ElorrietaNo ratings yet
- Flurosis Dental PDFDocument5 pagesFlurosis Dental PDFVanessa OrozcoNo ratings yet
- Micosis de Importancia OdontologícaDocument30 pagesMicosis de Importancia OdontologícaSarah VargasNo ratings yet
- Consideraciones A Tomar en Cuenta en El Manejo Odontológico Del Paciente Con Diabetes MellitusDocument6 pagesConsideraciones A Tomar en Cuenta en El Manejo Odontológico Del Paciente Con Diabetes Mellitusdopla11No ratings yet
- Patologia Lingual y EnciasDocument34 pagesPatologia Lingual y EnciasJhonathan MaldonadoNo ratings yet
- Complejo Dentino-PulparDocument5 pagesComplejo Dentino-Pulparsgarcia201082No ratings yet
- Resumen BiofilmDocument5 pagesResumen BiofilmCitlalli RguezNo ratings yet
- Bioquímica de La Saliva, Diente y PeriodontoDocument22 pagesBioquímica de La Saliva, Diente y PeriodontonormanNo ratings yet
- Manifestaciones Dentales de La Diabetes MellitusDocument10 pagesManifestaciones Dentales de La Diabetes MellitusAlejandra TerronNo ratings yet
- Neumonicosis FinalDocument31 pagesNeumonicosis FinalLuis Alfonso Asiain CanoNo ratings yet
- Irrigacion Del PeriodontoDocument12 pagesIrrigacion Del PeriodontoGaby Diaz de León50% (2)
- Características Del ADN y El Código Genético de Los HumanosDocument5 pagesCaracterísticas Del ADN y El Código Genético de Los Humanoscbuson100% (2)
- Manejo de Paciente SistemicoDocument37 pagesManejo de Paciente SistemicoPaúl SalazarNo ratings yet
- Glandulas SalivalesDocument39 pagesGlandulas SalivalesXiomara Oviedo RomeroNo ratings yet
- Micosis de La Mucosa OralDocument14 pagesMicosis de La Mucosa Oralvictor garcia100% (1)
- Paractica 1 InmunoDocument8 pagesParactica 1 InmunojordanNo ratings yet
- Embriología Del Sistema Estomatognátic1Document16 pagesEmbriología Del Sistema Estomatognátic1William FC UniversidadNo ratings yet
- Labio Leporino, GuiaDocument8 pagesLabio Leporino, GuiatigremedNo ratings yet
- Monografia Final Responsabilidad SocialDocument14 pagesMonografia Final Responsabilidad SocialGabriela CastañedaNo ratings yet
- ALOP 2017-1 FinalDocument67 pagesALOP 2017-1 Finaljavier100% (3)
- Citologia BasicaDocument33 pagesCitologia BasicaJhon Salas Mallqui0% (1)
- Tecnicas de Preparacion Biomecanica PDFDocument13 pagesTecnicas de Preparacion Biomecanica PDFWilliam La Madrid100% (1)
- Evaluación Del GustoDocument4 pagesEvaluación Del GustoLeonardo Ibañez Tejada100% (1)
- Genes HoxDocument12 pagesGenes HoxAndrea Cardenas FrancoNo ratings yet
- Bioquimica Del Diente 1Document28 pagesBioquimica Del Diente 1Yessica ChiuNo ratings yet
- Placa Bacteriana PDFDocument16 pagesPlaca Bacteriana PDFPara Tí Una Sonrisa100% (1)
- Halitosis, Tabaquismo y PeriodontitisDocument18 pagesHalitosis, Tabaquismo y PeriodontitisJessica OvandoNo ratings yet
- Diagnóstico y Tratamiento en EndodonciaDocument12 pagesDiagnóstico y Tratamiento en EndodonciaMarco ArmijosNo ratings yet
- Exploracion de La Cavidad OralDocument14 pagesExploracion de La Cavidad OralMarisol Mallcco AyalaNo ratings yet
- Saliva Cuadro 11Document2 pagesSaliva Cuadro 11Tefi MejíaNo ratings yet
- Tratamientos Pulpares en Dentición TemporalDocument41 pagesTratamientos Pulpares en Dentición TemporalEstefania Idrovo Yulan100% (1)
- Prueba 2 2019Document22 pagesPrueba 2 2019Joseph Cuaquera Castillo100% (1)
- Las Enfermedades Periodontales y Su Relación Con Enfermedades SistémicasDocument9 pagesLas Enfermedades Periodontales y Su Relación Con Enfermedades SistémicasRichard Omar Moreano MoreanoNo ratings yet
- Queratoquiste OdontogenicoDocument6 pagesQueratoquiste OdontogenicoLeonardo GilNo ratings yet
- Consideraciones en El Tratamiento Odontológico de Pacientes en Terapia Con BifosfonatosDocument12 pagesConsideraciones en El Tratamiento Odontológico de Pacientes en Terapia Con BifosfonatosSargento Rodrigo100% (2)
- Plasma Rico en Fibrina en PeriodontitisDocument88 pagesPlasma Rico en Fibrina en PeriodontitisLiliana Miranda ANo ratings yet
- 6618-Article Text-28300-1-10-20140206Document18 pages6618-Article Text-28300-1-10-20140206Magnolia CuelaNo ratings yet
- Manual de Referencia para Procedimientos en Odontopediatria Capitulo 19Document14 pagesManual de Referencia para Procedimientos en Odontopediatria Capitulo 19Andrés CeliNo ratings yet
- A Que Se Deben Las Caries en Los Pacientes DiabeticosDocument7 pagesA Que Se Deben Las Caries en Los Pacientes DiabeticosRiky Díaz CamposanoNo ratings yet
- Acidos GrabadoresDocument7 pagesAcidos GrabadoresRebeca SandovalNo ratings yet
- Enfermedad Periodontal Cultivo y Antibiograma DEFDocument23 pagesEnfermedad Periodontal Cultivo y Antibiograma DEFMichuu LascanoNo ratings yet
- Acetilcolina en Muscula Cardiaco PDFDocument10 pagesAcetilcolina en Muscula Cardiaco PDFGonzalo UribeNo ratings yet
- Morfofunción II Caso ClínicoDocument5 pagesMorfofunción II Caso ClínicoKaren Jessenia Gómez AlcivarNo ratings yet
- Dentición Temporal y MixtaDocument43 pagesDentición Temporal y MixtaSantiago AmelinesNo ratings yet
- Aislamiento AbsolutoDocument15 pagesAislamiento AbsolutoJosé Poveda0% (1)
- Sindrome de Boca Ardiente UASDDocument9 pagesSindrome de Boca Ardiente UASDPamela Mejia100% (1)
- Degradacion y Recuperacion de Las Bases Puricas y PirimidicasDocument10 pagesDegradacion y Recuperacion de Las Bases Puricas y PirimidicasBren Morales NegrellosNo ratings yet
- Luis Baeza Inteligencia Artificial Aplicada A La Odontología PDFDocument1 pageLuis Baeza Inteligencia Artificial Aplicada A La Odontología PDFLuis Ignacio Baeza GonzálezNo ratings yet
- Biología Molecular y Celular de Los Tejidos Periodontales Sanos y EnfermosDocument21 pagesBiología Molecular y Celular de Los Tejidos Periodontales Sanos y EnfermosAriel Gottlieb Palombo100% (2)
- Sofa 160905051938Document9 pagesSofa 160905051938marco antonio100% (1)
- Tratamiento de La Bolsa PeriodontalDocument18 pagesTratamiento de La Bolsa PeriodontalAnonymous DkgwveNo ratings yet
- Continuidad Anatómica 2Document4 pagesContinuidad Anatómica 2Diego PatiñoNo ratings yet
- Manchas Blancas OrtodonciaDocument7 pagesManchas Blancas Ortodonciakevin ramirez alonsoNo ratings yet
- Los Diferentes Tipos de Colgajos en Las Relaciones Quirúrgicas de La Tercera Molares ImpactadosDocument4 pagesLos Diferentes Tipos de Colgajos en Las Relaciones Quirúrgicas de La Tercera Molares ImpactadosDaniela DelgadoNo ratings yet
- AVULSIÓN PPT AlexDocument21 pagesAVULSIÓN PPT AlexJaime Alexis CANo ratings yet
- Transdisciplinariedad y Pensamiento ComplejoDocument1 pageTransdisciplinariedad y Pensamiento ComplejoasemiveNo ratings yet
- Análisis Artículos Leyes VenezolanasDocument9 pagesAnálisis Artículos Leyes VenezolanasasemiveNo ratings yet
- Análisis Escuela PunitivaDocument5 pagesAnálisis Escuela PunitivaasemiveNo ratings yet
- Artículo Orientación EducativaDocument4 pagesArtículo Orientación EducativaasemiveNo ratings yet
- TransdisciplinariedadDocument1 pageTransdisciplinariedadasemiveNo ratings yet
- Plan DidácticoDocument2 pagesPlan DidácticoasemiveNo ratings yet
- Proceso CivilDocument3 pagesProceso CivilasemiveNo ratings yet
- Ensayo Descripcion de GobiernoDocument3 pagesEnsayo Descripcion de GobiernoasemiveNo ratings yet
- Transdisciplinariedad y Pensamiento ComplejoDocument1 pageTransdisciplinariedad y Pensamiento ComplejoasemiveNo ratings yet
- Análisis Artículos Leyes VenezolanasDocument9 pagesAnálisis Artículos Leyes VenezolanasasemiveNo ratings yet
- Ensayo Herencia PoliticaDocument3 pagesEnsayo Herencia PoliticaasemiveNo ratings yet
- Cultura OrganizacionalDocument3 pagesCultura OrganizacionalasemiveNo ratings yet
- Análisis Escuela PunitivaDocument5 pagesAnálisis Escuela PunitivaasemiveNo ratings yet
- Referencia LaboralDocument1 pageReferencia LaboralasemiveNo ratings yet
- Delito Imposible DoxDocument4 pagesDelito Imposible DoxasemiveNo ratings yet
- Venezuela PetroleraDocument7 pagesVenezuela PetroleraasemiveNo ratings yet
- Artículo InvestigaciónDocument1 pageArtículo InvestigaciónasemiveNo ratings yet
- Análisis Artículos Leyes VenezolanasDocument9 pagesAnálisis Artículos Leyes VenezolanasasemiveNo ratings yet
- Análisis ArtículosDocument4 pagesAnálisis ArtículosasemiveNo ratings yet
- Análisis Comparativo Ley Tutelar Del MenorDocument5 pagesAnálisis Comparativo Ley Tutelar Del MenorasemiveNo ratings yet
- Conceptos DentalesDocument13 pagesConceptos DentalesasemiveNo ratings yet
- Educación A DistanciaDocument6 pagesEducación A DistanciaasemiveNo ratings yet
- Tríptico DPDocument2 pagesTríptico DPasemiveNo ratings yet
- Crisis ConstitucionalDocument3 pagesCrisis ConstitucionalasemiveNo ratings yet
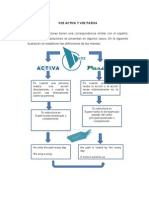
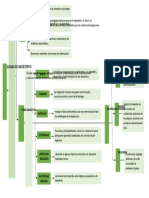
- Voz Activa y Voz PasivaDocument7 pagesVoz Activa y Voz PasivaasemiveNo ratings yet
- Planificación Del II Lapso - Lengua y Literatura - 1º U - L. B. José Félix RibasDocument3 pagesPlanificación Del II Lapso - Lengua y Literatura - 1º U - L. B. José Félix RibasasemiveNo ratings yet
- Desarrollo de La Gramtica A Proposito de La Comprensin y Produccin de Textos NarrativosDocument56 pagesDesarrollo de La Gramtica A Proposito de La Comprensin y Produccin de Textos NarrativosAlexis HernandezNo ratings yet
- Manual de Predicación BíblicaDocument50 pagesManual de Predicación BíblicaasemiveNo ratings yet
- Predicación - Una Decisión EquivocadaDocument2 pagesPredicación - Una Decisión EquivocadaasemiveNo ratings yet
- Enterogastrona Fisiología y Fisiopatología Pablo Gonzalez 201604136Document34 pagesEnterogastrona Fisiología y Fisiopatología Pablo Gonzalez 201604136Sebastián González100% (1)
- Cuestionario Sobre El Tema de Los Nutrientes CompleteDocument5 pagesCuestionario Sobre El Tema de Los Nutrientes Completejose100% (3)
- 10.tejido AdiposoDocument11 pages10.tejido AdiposoAlma Rosa OrtizNo ratings yet
- Nutrición de Las PlantasDocument6 pagesNutrición de Las PlantasMercedes Mendez RojasNo ratings yet
- Practica 12. Sds-PageDocument4 pagesPractica 12. Sds-PageMona MoralesNo ratings yet
- Examen Farmcologia FisioterapiaDocument2 pagesExamen Farmcologia FisioterapiaAlicia lópezNo ratings yet
- Bloque 2Document10 pagesBloque 2Miry RosalesNo ratings yet
- Síndromes de Inestabilidad Cromosómica Asociados ConDocument10 pagesSíndromes de Inestabilidad Cromosómica Asociados Conjessicarivera1994No ratings yet
- Meiosis y Gametogénesis para Quinto Grado de SecundariaDocument4 pagesMeiosis y Gametogénesis para Quinto Grado de SecundariaeduarNo ratings yet
- BiofisicaDocument3 pagesBiofisicaNancy FebrilletNo ratings yet
- Camu Camu Word ExposicionDocument7 pagesCamu Camu Word ExposicionAmabelia GuerrerosNo ratings yet
- Practica LevadurasDocument24 pagesPractica LevadurasCYNTHIA AZUCENA LIMA RUIZNo ratings yet
- Mapa Conceptual Del Aparato DigestivoDocument1 pageMapa Conceptual Del Aparato DigestivoAllison Arias67% (3)
- Análisis de Paternidad Usando CibertorioDocument2 pagesAnálisis de Paternidad Usando Cibertoriorepresentantebiologi50% (2)
- Tema 14. Sistema Calicreína-Cininonego-Cinina MODIFICADODocument6 pagesTema 14. Sistema Calicreína-Cininonego-Cinina MODIFICADOLuis MartinezNo ratings yet
- Preguntas Sobre Pruebas MolecularesDocument8 pagesPreguntas Sobre Pruebas MolecularesNats OCNo ratings yet
- Examenes de Sangre 01.07Document11 pagesExamenes de Sangre 01.07Roxana Matamala M.100% (1)
- Fase Oscura de La FotosintesisDocument12 pagesFase Oscura de La FotosintesisGerardo Rodrigo Alvarado AlfonsoNo ratings yet
- Cancer y Ciclo CelularDocument43 pagesCancer y Ciclo CelularOscar Manuel Monje CórdobaNo ratings yet
- Trabajo Colaborativo - 40 Tarea 2 FarmacologiaDocument13 pagesTrabajo Colaborativo - 40 Tarea 2 FarmacologiaLizeth HernándezNo ratings yet
- Resumen Teorias de EnvejecimientoDocument6 pagesResumen Teorias de EnvejecimientoLuis Alvarez100% (1)
- BI Sem7Document6 pagesBI Sem7Jose Edson MillaNo ratings yet
- Semana 13-BioquímicaDocument4 pagesSemana 13-BioquímicaLINDSAY CIANKINER CAMPOS SALDAÑANo ratings yet
- MIP-133 Pandemia 2020 Unidad 2Document65 pagesMIP-133 Pandemia 2020 Unidad 2maria Rambalde RamirezNo ratings yet
- Quimica 3Document3 pagesQuimica 3MaurenNo ratings yet
- Anual San Marcos - Biología Semana 02Document20 pagesAnual San Marcos - Biología Semana 02Giuliana CorcueraNo ratings yet
- Reaccion Antigeno - AnticuerpoDocument10 pagesReaccion Antigeno - AnticuerpoTJJeffersonNo ratings yet
- Taller 10. Desnaturalización de ProteinasDocument5 pagesTaller 10. Desnaturalización de ProteinasGabriel CaminoNo ratings yet
- Biología Celular Sesión 15 Organización General y Actividades Funcionales Del Núcleo InterfásicoDocument18 pagesBiología Celular Sesión 15 Organización General y Actividades Funcionales Del Núcleo InterfásicoRenataNo ratings yet