You might also like
- Koskeridou Et Al 2017 - Pliocene Grastropod Indicators GreeceDocument11 pagesKoskeridou Et Al 2017 - Pliocene Grastropod Indicators GreeceAndrew ButcherNo ratings yet
- National Geographic Little Kids March AprilDocument36 pagesNational Geographic Little Kids March AprilWillimSmith100% (8)
- Chapter 6 - Cell Parts!: AP Biology Cells Review!!! Name - DateDocument2 pagesChapter 6 - Cell Parts!: AP Biology Cells Review!!! Name - DateMaryam AllanaNo ratings yet
- Liraetal 2017Document12 pagesLiraetal 2017Claudeilton SantanaNo ratings yet
- DiatomDocument16 pagesDiatomRecto SutismaNo ratings yet
- New Records and Distribution of Marine Free-Living Nematodes in The Maldivian ArchipelagoDocument12 pagesNew Records and Distribution of Marine Free-Living Nematodes in The Maldivian ArchipelagoFrontiersNo ratings yet
- New Records of Marine Annelida (Polychaeta) in The Egyptian Coast, Eastern Mediterranean SeaDocument13 pagesNew Records of Marine Annelida (Polychaeta) in The Egyptian Coast, Eastern Mediterranean SeaSabrina Nur Fitri 2004125073No ratings yet
- tmpED44 TMPDocument14 pagestmpED44 TMPFrontiersNo ratings yet
- 1 s2.0 S0048969723018752 MainDocument13 pages1 s2.0 S0048969723018752 MainPedroHenriqueNo ratings yet
- A Preliminary Study of The Odonata of S'Albufera Natural Park, PDFDocument10 pagesA Preliminary Study of The Odonata of S'Albufera Natural Park, PDFOshLifin RucmanaNo ratings yet
- Cambios AnualesDocument22 pagesCambios AnualesFélix Armadá JuárezNo ratings yet
- A Predation Case of Anguis Graeca Bedriaga, 1881 (Squamata: Anguidae) by Lacerta Trilineata Bedriaga, 1886 (Squamata: Lacertidae) From Central GreeceDocument3 pagesA Predation Case of Anguis Graeca Bedriaga, 1881 (Squamata: Anguidae) by Lacerta Trilineata Bedriaga, 1886 (Squamata: Lacertidae) From Central GreeceRuben FernandezNo ratings yet
- fb06 PDFDocument29 pagesfb06 PDFtrivikramraoNo ratings yet
- 2.a Preliminary Study of The Odonata of SAlbufera Natural Park MallorcaDocument10 pages2.a Preliminary Study of The Odonata of SAlbufera Natural Park MallorcaAndreea MandacheNo ratings yet
- 2013 - Phylogenetic Identification of Marine BacteriaDocument10 pages2013 - Phylogenetic Identification of Marine BacteriaMarcus Adonai Castro da SilvaNo ratings yet
- Dalyan 2014Document2 pagesDalyan 2014Fadhli LatuconsinaNo ratings yet
- Integrative Redescription of A Common Arctic Water Bear Pilatobius Recamieri (Richters, 1911)Document14 pagesIntegrative Redescription of A Common Arctic Water Bear Pilatobius Recamieri (Richters, 1911)HagarNo ratings yet
- Ambroso Et Al 2016. Ophiuroid Biodiversity Patterns Along The Antarctic PeninsulaDocument16 pagesAmbroso Et Al 2016. Ophiuroid Biodiversity Patterns Along The Antarctic Peninsulabyhz7wwdm4No ratings yet
- Guidetti 2013Document4 pagesGuidetti 2013Fadhli LatuconsinaNo ratings yet
- Distribution and Ecology of Non Marine oDocument15 pagesDistribution and Ecology of Non Marine oJoseph HOTEKPONo ratings yet
- Anthropogenic Stressors, ENSODocument13 pagesAnthropogenic Stressors, ENSOCaro301079No ratings yet
- Potential Roles of Labyrinthula Spp. in Global Seagrass Population DeclinesDocument11 pagesPotential Roles of Labyrinthula Spp. in Global Seagrass Population DeclinesJose CejudoNo ratings yet
- Peran Ekologi Bulu Babi Dalam KomunitasDocument8 pagesPeran Ekologi Bulu Babi Dalam Komunitasenago moteNo ratings yet
- Ecology and Distribution of Aulacoseira Species Bacillariophyta in Tropical Reservoirs From BrazilDocument18 pagesEcology and Distribution of Aulacoseira Species Bacillariophyta in Tropical Reservoirs From BrazilPaola JiradoNo ratings yet
- 2019-Assessment On Recruitment Density of Branching Corals Montipora Digitata (Scleractinia Acroporidae) in Talairi Island, Gulf of MannarDocument4 pages2019-Assessment On Recruitment Density of Branching Corals Montipora Digitata (Scleractinia Acroporidae) in Talairi Island, Gulf of MannarRameshNo ratings yet
- Oray 2015Document3 pagesOray 2015Hector RodriguezNo ratings yet
- 44023-Article Text-138036-150338-10-20181227Document11 pages44023-Article Text-138036-150338-10-20181227Lara Caroline Carvalho RezendeNo ratings yet
- Jellyfish Aggregations and Leatherback Turtle Foraging Patterns in A Temperate Coastal EnvironmentDocument6 pagesJellyfish Aggregations and Leatherback Turtle Foraging Patterns in A Temperate Coastal EnvironmentBengt HörbergNo ratings yet
- An Overview of The Dragonflies and Damselflies of The Maltese Islands (Central Mediterranean) (Odonata)Document123 pagesAn Overview of The Dragonflies and Damselflies of The Maltese Islands (Central Mediterranean) (Odonata)Ram LeeNo ratings yet
- Nesterova BS Phyto 2008Document34 pagesNesterova BS Phyto 2008Metehan KaraNo ratings yet
- Yap Et Al 2023 Taxonomy and Molecular Phylogeny of The Sea Anemone Macrodactyla With A Description of A New Species From SingaporeDocument25 pagesYap Et Al 2023 Taxonomy and Molecular Phylogeny of The Sea Anemone Macrodactyla With A Description of A New Species From SingaporenickYNo ratings yet
- EJABF-Volume 24-Issue 6 - Page 435-457Document23 pagesEJABF-Volume 24-Issue 6 - Page 435-457SADEQNo ratings yet
- Jurnal Sebaran BivalviaDocument11 pagesJurnal Sebaran Bivalviafella suffa azzahraNo ratings yet
- Larsson Et Al 2018Document12 pagesLarsson Et Al 2018Nadia Valeria HerreraNo ratings yet
- 1194-Full Length Article-3380-1-10-20220618Document14 pages1194-Full Length Article-3380-1-10-20220618France Lee ManobanNo ratings yet
- Pimiento Et Al-2016Document11 pagesPimiento Et Al-2016star warsNo ratings yet
- Diversity of PlanktonDocument10 pagesDiversity of Planktonsara unbeknaNo ratings yet
- Journal Homepage: - : IntroductionDocument12 pagesJournal Homepage: - : IntroductionIJAR JOURNALNo ratings yet
- Sea Star Prop.Document10 pagesSea Star Prop.Mia JaiNo ratings yet
- Fuentes Et Al. 2011 - Rhizostoma Pulmo Life CycleDocument21 pagesFuentes Et Al. 2011 - Rhizostoma Pulmo Life CycleAmrutha AravindNo ratings yet
- 1 s2.0 S003515982030043X MainDocument11 pages1 s2.0 S003515982030043X MainEleonoraNo ratings yet
- tmp909F TMPDocument12 pagestmp909F TMPFrontiersNo ratings yet
- NoteDocument8 pagesNoteشهرول عفيفيNo ratings yet
- Biogeography of Jellyfish in The North Atlantic, by Traditional and Genomic MethodsDocument19 pagesBiogeography of Jellyfish in The North Atlantic, by Traditional and Genomic MethodsYovan BlancoNo ratings yet
- 1 s2.0 S1517838217307815 MainDocument8 pages1 s2.0 S1517838217307815 MainMuhamad AliNo ratings yet
- Ecological Understanding For Fishery Management Condition and Growth of Anchovy Late LarvaeDocument9 pagesEcological Understanding For Fishery Management Condition and Growth of Anchovy Late LarvaeSolansh Reyes MendoNo ratings yet
- This Is The Published VersionDocument7 pagesThis Is The Published VersionAlínio JoséNo ratings yet
- Woo, 2013Document6 pagesWoo, 2013CroatoanNo ratings yet
- Souter Et Al 2009 BMC EcologyDocument13 pagesSouter Et Al 2009 BMC EcologyBhavna RamkissoonNo ratings yet
- Euphausia Sibogae: Research ArticleDocument12 pagesEuphausia Sibogae: Research ArticlelijojhonNo ratings yet
- Reprodução Wiederheckeretal2002Document23 pagesReprodução Wiederheckeretal2002Cristine PratesNo ratings yet
- Paleoenvironment and Paleoecology Based On Recent Benthic Foraminifera in Cikadal Beach, Ciletuh Geopark, West Java, IndonesiaDocument7 pagesPaleoenvironment and Paleoecology Based On Recent Benthic Foraminifera in Cikadal Beach, Ciletuh Geopark, West Java, IndonesiaPanjiNo ratings yet
- World Distribution, Diversity and Endemism of Aquatic MacrophytesDocument16 pagesWorld Distribution, Diversity and Endemism of Aquatic MacrophytesJonathan Urrutia EstradaNo ratings yet
- Robinsonetal 2023Document15 pagesRobinsonetal 2023Elmo KochNo ratings yet
- Environmental Microbiology 64 (10), 3748-53.: ReferencesDocument5 pagesEnvironmental Microbiology 64 (10), 3748-53.: ReferencesKristelle AbadNo ratings yet
- Wood Etal 1999Document21 pagesWood Etal 1999Mateus Felipe Miranda BatistaNo ratings yet
- Macrobenthos of Meghna River Estuary, BangaldeshDocument7 pagesMacrobenthos of Meghna River Estuary, BangaldeshMohammad Belal HossainNo ratings yet
- Diversity of Phytoplankton in Coastal Water of Kuantan, Pahang, MalaysiaDocument9 pagesDiversity of Phytoplankton in Coastal Water of Kuantan, Pahang, MalaysianabilaNo ratings yet
- Coral Disease Prevalence and Coral Health in The Wakatobi Marine Park, South-East Sulawesi, IndonesiaDocument13 pagesCoral Disease Prevalence and Coral Health in The Wakatobi Marine Park, South-East Sulawesi, IndonesiaHendra KaaNo ratings yet
- First Report of The Larva of A Vulnerable Damselfly in Nigeria, With Some Ecological NotesDocument7 pagesFirst Report of The Larva of A Vulnerable Damselfly in Nigeria, With Some Ecological Notesabiodunblaise15No ratings yet
- Use Only: Hard Times For Catadromous Fish: The Case of The European Eel Anguilla Anguilla (L. 1758)Document18 pagesUse Only: Hard Times For Catadromous Fish: The Case of The European Eel Anguilla Anguilla (L. 1758)philosophy-thoughtNo ratings yet
- Marine MacroecologyFrom EverandMarine MacroecologyJon D. WitmanNo ratings yet
- Jiaa 405Document5 pagesJiaa 405spin_echoNo ratings yet
- Mouse Anti-Flavivirus Group Antigen Monoclonal AntibodyDocument1 pageMouse Anti-Flavivirus Group Antigen Monoclonal Antibodyspin_echoNo ratings yet
- Martin Palus Tick-Borne Encephalitis - From PathogenesisDocument213 pagesMartin Palus Tick-Borne Encephalitis - From Pathogenesisspin_echoNo ratings yet
- Lima Barbero2019Document10 pagesLima Barbero2019spin_echoNo ratings yet
- 751 FullDocument6 pages751 Fullspin_echoNo ratings yet
- Plasma Donors Neutralisation AssayDocument6 pagesPlasma Donors Neutralisation Assayspin_echoNo ratings yet
- Online First: Full Research ArticleDocument14 pagesOnline First: Full Research Articlespin_echoNo ratings yet
- HLA - 2021 - de Marco - HLA A Homozygosis Is Associated With Susceptibility To COVID 19Document10 pagesHLA - 2021 - de Marco - HLA A Homozygosis Is Associated With Susceptibility To COVID 19spin_echoNo ratings yet
- Evaluation of Antivirals Against Tick-Borne Encephalitis Virus in Organotypic Brain Slices of Rat CerebellumDocument13 pagesEvaluation of Antivirals Against Tick-Borne Encephalitis Virus in Organotypic Brain Slices of Rat Cerebellumspin_echoNo ratings yet
- Correlation Between ELISA, Hemagglutination Inhibition, and Neutralization Tests After Vaccination Against Tick-Borne EncephalitisDocument6 pagesCorrelation Between ELISA, Hemagglutination Inhibition, and Neutralization Tests After Vaccination Against Tick-Borne Encephalitisspin_echoNo ratings yet
- First Detection of Spotted Fever Group Rickettsiae in Ticks in SerbiaDocument6 pagesFirst Detection of Spotted Fever Group Rickettsiae in Ticks in Serbiaspin_echoNo ratings yet
- PPSRPS ISO 9001-2015++ (S)Document40 pagesPPSRPS ISO 9001-2015++ (S)spin_echoNo ratings yet
- Induction, Purification and Characterization of An Antibacterial Peptide Scolopendrin I From The Venom of Centipede Scolopendra Subspinipes MutilansDocument6 pagesInduction, Purification and Characterization of An Antibacterial Peptide Scolopendrin I From The Venom of Centipede Scolopendra Subspinipes Mutilansspin_echoNo ratings yet
- Hasan2004 PDFDocument2 pagesHasan2004 PDFspin_echoNo ratings yet
- Sumner 2011Document12 pagesSumner 2011spin_echoNo ratings yet
- Condyle ApproachDocument5 pagesCondyle Approachspin_echoNo ratings yet
- Adjuvants Designed For Veterinary and Human Vaccines - 2001 - Vaccine PDFDocument7 pagesAdjuvants Designed For Veterinary and Human Vaccines - 2001 - Vaccine PDFDeboraXiningNo ratings yet
- Veraldi 2010Document2 pagesVeraldi 2010spin_echoNo ratings yet
- Harnessing Bene Fit From Targeting Tumor Associated Carbohydrate AntigensDocument9 pagesHarnessing Bene Fit From Targeting Tumor Associated Carbohydrate Antigensspin_echoNo ratings yet
- Morphometric Analysis Related To The TranscondylarDocument6 pagesMorphometric Analysis Related To The Transcondylarspin_echoNo ratings yet
- Unilateral, Double Hypoglossal Nerves Leaving The Cranial Cavity Through Two HypoglossalDocument2 pagesUnilateral, Double Hypoglossal Nerves Leaving The Cranial Cavity Through Two Hypoglossalspin_echoNo ratings yet
- Morphometry of The Hypoglossal Canal, OccipitalDocument5 pagesMorphometry of The Hypoglossal Canal, Occipitalspin_echoNo ratings yet
- The Hypoglossal CanalDocument4 pagesThe Hypoglossal Canalspin_echoNo ratings yet
- Variations in The BridgingTraitof TheDocument6 pagesVariations in The BridgingTraitof Thespin_echoNo ratings yet
- Morphometric Analysis of The Foramen Magnum: An Anatomic StudyDocument4 pagesMorphometric Analysis of The Foramen Magnum: An Anatomic Studyspin_echoNo ratings yet
- The Hypoglossal Artery and Hypoglossal CanalDocument6 pagesThe Hypoglossal Artery and Hypoglossal Canalspin_echoNo ratings yet
- Median (Third) Occipital CondyleDocument4 pagesMedian (Third) Occipital Condylespin_echoNo ratings yet
- Variations in The BridgingTraitof TheDocument6 pagesVariations in The BridgingTraitof Thespin_echoNo ratings yet
- Occipital Condyle SyndromeDocument5 pagesOccipital Condyle Syndromespin_echoNo ratings yet
- Morphology (Biology)Document7 pagesMorphology (Biology)BrianNo ratings yet
- Botany ReviewerDocument14 pagesBotany ReviewerTherese EspinosaNo ratings yet
- Processes of EvolutionDocument88 pagesProcesses of EvolutionDiane HeartphiliaNo ratings yet
- Helpful Insects, Spiders and PathogensDocument138 pagesHelpful Insects, Spiders and PathogensAnonymous HXLczq3100% (1)
- BiologistDocument51 pagesBiologistJean CabigaoNo ratings yet
- Homo FloresiensisDocument14 pagesHomo Floresiensissifa100% (1)
- Invading AntsDocument12 pagesInvading Antshamidreza mohseniNo ratings yet
- Sound 3-4 QR Usingechoestosee - HighDocument1 pageSound 3-4 QR Usingechoestosee - HighLola LolaNo ratings yet
- Hunt (2008) - Anniella PDFDocument11 pagesHunt (2008) - Anniella PDFKlaus FischerNo ratings yet
- ALUMBHEGDocument21 pagesALUMBHEGabed9631No ratings yet
- The Wreck of The Zanzibar ResourcesDocument18 pagesThe Wreck of The Zanzibar ResourcesMonica Scaltrito100% (1)
- Evidence of Human EvolutionDocument6 pagesEvidence of Human EvolutionSonia Sánchez BarrancoNo ratings yet
- Quiz - 6625 - Cambridge Starters Test 5 - Reading WritingDocument7 pagesQuiz - 6625 - Cambridge Starters Test 5 - Reading WritingDang Huyen LinhNo ratings yet
- Molecular Biology of Cellulose Production in Bacteria: Ute RömlingDocument8 pagesMolecular Biology of Cellulose Production in Bacteria: Ute Römlingrizanda93No ratings yet
- EvolutionDocument25 pagesEvolutionapi-264004571No ratings yet
- Growth PredictionDocument64 pagesGrowth PredictionMothi KrishnaNo ratings yet
- Bio 11 UPD Third Lec Exam ReviewerDocument111 pagesBio 11 UPD Third Lec Exam ReviewerMedjugorje Beltran DalanNo ratings yet
- Making A Boneless BangusDocument2 pagesMaking A Boneless BangusRommel P. AbasNo ratings yet
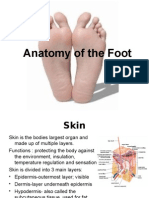
- Anatomy FootDocument22 pagesAnatomy Footazis aimaduddinNo ratings yet
- Medical Parasitology - FullDocument30 pagesMedical Parasitology - FullJesse Osborn100% (2)
- Class XI English Hornbill The Laburnum Top NotesDocument5 pagesClass XI English Hornbill The Laburnum Top NotesParvani Patankar100% (1)
- Use of Insects in Forensic ScienceDocument3 pagesUse of Insects in Forensic ScienceEmanuel Ruiz RodriguezNo ratings yet
- Understanding Samhita With Drushtanta W.S.R OrnithologyDocument5 pagesUnderstanding Samhita With Drushtanta W.S.R OrnithologyAna SharmaNo ratings yet
- WK Sheet Connect Plus Model AnsweDocument5 pagesWK Sheet Connect Plus Model Answemarinasabry123456No ratings yet
- The Ultra Structure of Prokaryotic-Eukaryotic CellDocument18 pagesThe Ultra Structure of Prokaryotic-Eukaryotic CellNicholas Music-SchoolNo ratings yet
- BeautyDocument2 pagesBeautyPraghashrajaNo ratings yet
- Parasitology Chart 211Document20 pagesParasitology Chart 211Sanjay Kumar100% (1)
- Virtual Field Trip Report To Kopf Family Reservation July 2021Document41 pagesVirtual Field Trip Report To Kopf Family Reservation July 2021Western Cuyahoga AudubonNo ratings yet