You might also like
- Práctica 17. NucleoloDocument5 pagesPráctica 17. NucleoloAndrea TovarNo ratings yet
- Tarea Resumen El Nucleolo Como Un Regulador Del Envejecimiento CelularDocument4 pagesTarea Resumen El Nucleolo Como Un Regulador Del Envejecimiento Celularkatherin PCNo ratings yet
- Taller Sobre Lectura Crítica - Ana Milena RuizDocument4 pagesTaller Sobre Lectura Crítica - Ana Milena RuizAna Milena Ruiz RomeroNo ratings yet
- Nucleo y NucleoloDocument3 pagesNucleo y NucleoloFelix Gutierrez100% (1)
- NucleologenesisDocument48 pagesNucleologenesisRodolfo Castillo50% (2)
- Informe de Exposición NúcleoDocument14 pagesInforme de Exposición Núcleomafer salazarNo ratings yet
- INTRODUCCION3333Document2 pagesINTRODUCCION3333JorgeNo ratings yet
- Nucleo y NucleoloDocument3 pagesNucleo y NucleoloJuan Carlos Vázquez92% (13)
- El Nucléolo enDocument29 pagesEl Nucléolo enFredy Alberto CabasNo ratings yet
- 11 - Arquitectura NuclearDocument9 pages11 - Arquitectura NuclearJhon RVNo ratings yet
- Informe de BioCel - NucleoloDocument7 pagesInforme de BioCel - NucleoloJuan Llontop SolanoNo ratings yet
- El Nucleolo en EspanolDocument7 pagesEl Nucleolo en Espanolmateo charrisNo ratings yet
- Nucleo ArquitecturaDocument8 pagesNucleo Arquitecturaarcadia_hbNo ratings yet
- Unidad 9 ReplicDocument29 pagesUnidad 9 ReplicLUDMILA FERNÁNDEZNo ratings yet
- Unidad 09 ReplicacionDocument12 pagesUnidad 09 ReplicacionMicaela FragaNo ratings yet
- El Nucleolo (Resumen)Document6 pagesEl Nucleolo (Resumen)Angel Maqquerhua PereyraNo ratings yet
- Los Límites Entre La Histología y La Bioquímica Observando Al Núcleo CelularDocument16 pagesLos Límites Entre La Histología y La Bioquímica Observando Al Núcleo Celularedson floresNo ratings yet
- Articulos CientificosDocument4 pagesArticulos CientificosAlejandra Arcos GuerreroNo ratings yet
- El NucleoDocument10 pagesEl NucleoSmith Rels100% (1)
- Nucleo InterfasticoDocument18 pagesNucleo InterfasticoAnahi Alban100% (1)
- Trabajo Práctico #2.Document23 pagesTrabajo Práctico #2.Juan Cruz FARFANNo ratings yet
- Triptico Celula BiologiaDocument3 pagesTriptico Celula BiologiaMIRNA75% (4)
- La Frontera Genetica Molecular de La EnfermedadDocument526 pagesLa Frontera Genetica Molecular de La EnfermedadDeivy Horta Tkd0% (1)
- Info. Importante Nucleo y NucleoloDocument4 pagesInfo. Importante Nucleo y NucleoloAlejandraNo ratings yet
- Nucleo InterfasicoDocument16 pagesNucleo InterfasicoVane PonsNo ratings yet
- La Frontera Genetica - MolecularDocument526 pagesLa Frontera Genetica - MolecularFrancis Vallecillo PadillaNo ratings yet
- El NucleoloDocument6 pagesEl NucleoloHanna ZavalaNo ratings yet
- Biologia Trabajo CelulaDocument49 pagesBiologia Trabajo CelulaKARLA BAUTISTANo ratings yet
- El Daño Tisular y La Senescencia Proporcionan Señales Críticas para La Reprogramación Celular in VivoDocument24 pagesEl Daño Tisular y La Senescencia Proporcionan Señales Críticas para La Reprogramación Celular in VivoManuel David Fuentes GarciaNo ratings yet
- NUCLEODocument15 pagesNUCLEOEsmi Esmi TexotlticitlNo ratings yet
- Núcleo CelularDocument32 pagesNúcleo CelularAlisson PonceNo ratings yet
- Clase Estructura Del NucleoDocument90 pagesClase Estructura Del Nucleohghbkgykmgxyh100% (1)
- Genética SeminarioDocument7 pagesGenética SeminarioMayerling V. RiosNo ratings yet
- Resumen Nucleo EYFAMDocument8 pagesResumen Nucleo EYFAMBbm NosbornNo ratings yet
- Semana 7 - TeoriaDocument23 pagesSemana 7 - TeoriajoselinNo ratings yet
- Informe de Biologia Nucleo CelularDocument11 pagesInforme de Biologia Nucleo CelularelmerNo ratings yet
- Informe de Biologia Nucleo CelularDocument11 pagesInforme de Biologia Nucleo CelularelmerNo ratings yet
- El Núcleo CelularDocument3 pagesEl Núcleo CelularGinger AvilesNo ratings yet
- Guía TP 5Document16 pagesGuía TP 5Dani Morales DellatorreNo ratings yet
- Que Es El Jugo NuclearDocument6 pagesQue Es El Jugo NuclearPaúl Fernando100% (1)
- Atlas Celula 04 NucleoDocument20 pagesAtlas Celula 04 Nucleomanuel rodriguesNo ratings yet
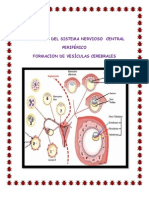
- Sistema Nervioso y Vesiculas CerebralesDocument35 pagesSistema Nervioso y Vesiculas Cerebraleslorietarosemary100% (1)
- Nucleo CelularDocument8 pagesNucleo CelularJHARIM NOELY AUCCAPIÑA GUILLENNo ratings yet
- Organización NuclearDocument6 pagesOrganización NuclearLuciana ColqueNo ratings yet
- Núcleo CelularDocument6 pagesNúcleo CelularBetzy Valery Poma PerezNo ratings yet
- NucleoloDocument2 pagesNucleoloMorales Tecalero Donaldo100% (1)
- Celulas Del Sistema NerviosoDocument12 pagesCelulas Del Sistema NerviosoSamayraNo ratings yet
- NucleoloDocument15 pagesNucleololeNo ratings yet
- División Celular y OncogenesDocument2 pagesDivisión Celular y OncogenesAna Martínez TorresNo ratings yet
- PANIAGUA Biologia Celular 3a Edicion - Estructura y Expresión Genética - ComprimidoDocument59 pagesPANIAGUA Biologia Celular 3a Edicion - Estructura y Expresión Genética - ComprimidosuldenyNo ratings yet
- Tema 8. NucleoDocument17 pagesTema 8. NucleoVanessa PeñaNo ratings yet
- Trabajo Practico #1. Biologia NucleoDocument17 pagesTrabajo Practico #1. Biologia NucleoMarcela BarriosNo ratings yet
- Tema 10 - El Núcleo - Reproducción CelularDocument23 pagesTema 10 - El Núcleo - Reproducción Celularcandelaropero2006No ratings yet
- Semana 7 Teoria - Nucleo CelularDocument17 pagesSemana 7 Teoria - Nucleo CelularEliza huamaniNo ratings yet
- Articulo Martinez Miguel JenniferDocument47 pagesArticulo Martinez Miguel Jenniferkatherin PCNo ratings yet
- Nucleo CelularDocument3 pagesNucleo CelularMarleni RobayoNo ratings yet
- Ultra Estructura Del Núcleo de La CélulaDocument13 pagesUltra Estructura Del Núcleo de La CélulaEduardo Yamunaqué CastroNo ratings yet
- Rejuvenecer Con El Plasma Sanguíneo De Los JóvenesFrom EverandRejuvenecer Con El Plasma Sanguíneo De Los JóvenesRating: 5 out of 5 stars5/5 (1)
- U1 Acto Moral 2020Document23 pagesU1 Acto Moral 2020Randolfo MaximilianoNo ratings yet
- Mapa Conceptual de Bacteriolog A y Virolog A M Dicas-5Document2 pagesMapa Conceptual de Bacteriolog A y Virolog A M Dicas-5Randolfo MaximilianoNo ratings yet
- Virologia - Conceptos BasicosDocument38 pagesVirologia - Conceptos BasicosRandolfo MaximilianoNo ratings yet
- Relieve de Amer - Clase 5Document2 pagesRelieve de Amer - Clase 5Randolfo MaximilianoNo ratings yet
- Teoria de AproximacionesDocument3 pagesTeoria de AproximacionesRandolfo MaximilianoNo ratings yet
- Guias de TPDocument33 pagesGuias de TPRandolfo MaximilianoNo ratings yet
- Teorico Practico 1 BioeticaDocument36 pagesTeorico Practico 1 BioeticaRandolfo MaximilianoNo ratings yet
- Teoria de AproximacionesDocument3 pagesTeoria de AproximacionesRandolfo MaximilianoNo ratings yet
- Guión Litúrgico de Solemne Eucaristía en Honor A San José (Recuperado Automáticamente)Document2 pagesGuión Litúrgico de Solemne Eucaristía en Honor A San José (Recuperado Automáticamente)Randolfo Maximiliano100% (2)
- Fermentación de AzucaresDocument2 pagesFermentación de AzucaresRandolfo MaximilianoNo ratings yet
- Factores Que Favorecieron El Estudio de La BioéticaDocument4 pagesFactores Que Favorecieron El Estudio de La BioéticaRandolfo MaximilianoNo ratings yet
- Belen Alejandra Sirvan Baglietto PDFDocument1 pageBelen Alejandra Sirvan Baglietto PDFRandolfo MaximilianoNo ratings yet
- Estatuto Docente PDFDocument144 pagesEstatuto Docente PDFRandolfo MaximilianoNo ratings yet
- Guia de Actividades Ingenieria GeneticaDocument9 pagesGuia de Actividades Ingenieria GeneticaRandolfo MaximilianoNo ratings yet
- Belen Alejandra Sirvan BagliettoDocument1 pageBelen Alejandra Sirvan BagliettoRandolfo MaximilianoNo ratings yet
- Leyes y NormasDocument6 pagesLeyes y NormasRandolfo MaximilianoNo ratings yet
- Leyes y NormasDocument6 pagesLeyes y NormasRandolfo MaximilianoNo ratings yet
- P53Document4 pagesP53CARLOS JESUS GARCIA HERNANDEZNo ratings yet
- Estabilización de Las Interacciones Proteína-Proteína en El Descubrimiento de FármacosDocument57 pagesEstabilización de Las Interacciones Proteína-Proteína en El Descubrimiento de FármacosDavid VasquezNo ratings yet
- Osteosarcoma: A. ResumenDocument33 pagesOsteosarcoma: A. ResumenHOSPITAL GENERAL DE LIBRESNo ratings yet
- Unidad 8 - NeoplasiasDocument25 pagesUnidad 8 - NeoplasiasLourdes VelazquezNo ratings yet
- Cuerpos de CajalDocument233 pagesCuerpos de CajalJuan Cruz FARFANNo ratings yet
- Arrest in AsDocument8 pagesArrest in AsWalter RamirezNo ratings yet
- Genes Estabilizadores y Su Relación Con Algúna Patogenisidad, Así Como Su Mecanismos de Señalización Al Cual Están Vinculados.Document5 pagesGenes Estabilizadores y Su Relación Con Algúna Patogenisidad, Así Como Su Mecanismos de Señalización Al Cual Están Vinculados.Daniel AscencioNo ratings yet
- Tema 33 - Genes Supresores de Tumores SARADocument10 pagesTema 33 - Genes Supresores de Tumores SARAJuan Manuel Morón OcañaNo ratings yet
- PICHER - El Nucléolo Como Sensor de Estrés Oxidativo. Implicaciones en Patologías Progeroides.Document54 pagesPICHER - El Nucléolo Como Sensor de Estrés Oxidativo. Implicaciones en Patologías Progeroides.katherin PCNo ratings yet
- 05 - Clase AntigenosDocument95 pages05 - Clase AntigenosdannymalcacamposNo ratings yet
- p53 en El Proceso de Reparación de Daño de AdnDocument5 pagesp53 en El Proceso de Reparación de Daño de AdnKaren GonzalesNo ratings yet
- 1 Ciclo Celular y CáncerDocument6 pages1 Ciclo Celular y CáncerLISETH KATHERINE RAMIREZ GARCIANo ratings yet
- B-Arrestin Mediated Receptor Trafficking and Signal TraductionDocument13 pagesB-Arrestin Mediated Receptor Trafficking and Signal TraductionSILVIA FERNANDA CASTILLO GOYENECHENo ratings yet
- P53 y MamaDocument33 pagesP53 y MamaNestor lorenzoNo ratings yet
- Sarcomas RetroperitonealesDocument58 pagesSarcomas RetroperitonealesSara MunozNo ratings yet
- Tema11 - RESPUESTA CELULAR AL DAÑO DEL ADNDocument34 pagesTema11 - RESPUESTA CELULAR AL DAÑO DEL ADNLidii monstersNo ratings yet
- Boletín de La Academia de Medicina Chilena. 2007Document246 pagesBoletín de La Academia de Medicina Chilena. 2007Camila Cisternas FloresNo ratings yet
- Adenomas y Adenocarcinoma de GL HepatoidesDocument32 pagesAdenomas y Adenocarcinoma de GL HepatoidesAlfredo MoralesNo ratings yet
- Taller 1 y 2 Biologia Celular y MolecularDocument53 pagesTaller 1 y 2 Biologia Celular y MolecularVanessa NúñezNo ratings yet
- Radiación Ultravioleta Cancer Piel P53Document8 pagesRadiación Ultravioleta Cancer Piel P53AlexRodrigoNo ratings yet
- Bioinorgánica Del ZNDocument10 pagesBioinorgánica Del ZNJesus27No ratings yet
- Regulación Ciclo Celular y El CancerDocument5 pagesRegulación Ciclo Celular y El CancerSantos Olav Godoy MoránNo ratings yet