You might also like
- Remedios Caseros - Ingrid PegueroDocument657 pagesRemedios Caseros - Ingrid PegueroJosé Héctor López Quintero100% (2)
- Embarazo y epigenética: el futuro de tu hijo es hoy…: Método GestacciónFrom EverandEmbarazo y epigenética: el futuro de tu hijo es hoy…: Método GestacciónNo ratings yet
- Transforma Tu CuerpoDocument31 pagesTransforma Tu CuerpoDiana Cristina Mejia100% (1)
- Ebook - Guia para Perder GrasaDocument24 pagesEbook - Guia para Perder GrasaMariano GuerreraNo ratings yet
- Desarrollo cognitivo en niños prematuros y de bajo peso al nacerFrom EverandDesarrollo cognitivo en niños prematuros y de bajo peso al nacerNo ratings yet
- Dieta Cetogénica Mediterránea EspañolaDocument142 pagesDieta Cetogénica Mediterránea Españolawinter100% (8)
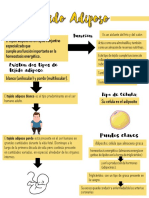
- Histologia Guia Del Tejido AdiposoDocument8 pagesHistologia Guia Del Tejido AdiposoOriana Sánchez100% (3)
- Crecimiento y Desarrollo Del EscolarDocument17 pagesCrecimiento y Desarrollo Del EscolarAnBCNo ratings yet
- Resultados maternos y fetales de la actividad física durante el embarazoFrom EverandResultados maternos y fetales de la actividad física durante el embarazoRating: 3 out of 5 stars3/5 (2)
- Manual de Crecimiento y Desarrollo Del NiñoDocument53 pagesManual de Crecimiento y Desarrollo Del NiñoSolanaHalogeno100% (2)
- Dossier Bioquímica de La Obesidad Des2016Document24 pagesDossier Bioquímica de La Obesidad Des2016Tina Monroy100% (1)
- Crecimiento y Desarrollo FetalDocument39 pagesCrecimiento y Desarrollo Fetalangela strzyzowskiNo ratings yet
- Tesis Factores Emocionales y Ambientales Que Predisponen ObesidadDocument127 pagesTesis Factores Emocionales y Ambientales Que Predisponen ObesidadIsabelEsnaolaNo ratings yet
- Adiccion A CarbohidratosDocument23 pagesAdiccion A CarbohidratosOsiris MissyNo ratings yet
- Cuidados en Trastornos Conducta AlimentariaDocument197 pagesCuidados en Trastornos Conducta Alimentariawww.ATENSALUD.comNo ratings yet
- Integración en El Metabolismo Del Ser Humano 01, Bioquímica Médica Universidad Científica Del SurDocument57 pagesIntegración en El Metabolismo Del Ser Humano 01, Bioquímica Médica Universidad Científica Del SurEnzon BritoNo ratings yet
- Fisiologa Del Crecimiento FetalDocument11 pagesFisiologa Del Crecimiento FetalROSMEL VLADIMIR FABIAN LLAJARUNANo ratings yet
- 2 Fisiología Del Crecimiento FetalDocument9 pages2 Fisiología Del Crecimiento FetalSarahí SosaNo ratings yet
- 2do Parcial StephanyaDocument30 pages2do Parcial StephanyaFanny GonzalezNo ratings yet
- Crecimiento Restringido Del FetoDocument15 pagesCrecimiento Restringido Del FetoMatias Israel CandoNo ratings yet
- Primer Expo Tercer GrupoDocument15 pagesPrimer Expo Tercer GrupoALLISON MICHAELLE VELEZ VILLAONo ratings yet
- 4-Chavira-Suarez-2017HIPOTESIS BARKERDocument8 pages4-Chavira-Suarez-2017HIPOTESIS BARKERsixtoNo ratings yet
- ASDocument19 pagesASRene Alexis Rodriguez JoaquinNo ratings yet
- Clase 4Document50 pagesClase 4Maricielo Samame LauraNo ratings yet
- Desarrollo Embriofetal Normal y PatológicoDocument15 pagesDesarrollo Embriofetal Normal y PatológicoDavid GuzmanNo ratings yet
- Exposicion Crecimiento y DesarrolloDocument51 pagesExposicion Crecimiento y DesarrolloGianfranco BuvoliNo ratings yet
- Capítulo 3 y 4Document16 pagesCapítulo 3 y 4mdmcenglfNo ratings yet
- Crecimienot y Desarrollo Del AdolescenteDocument30 pagesCrecimienot y Desarrollo Del AdolescenteMaydelin GuzmánNo ratings yet
- Clase Crecimiento y DesarrolloDocument52 pagesClase Crecimiento y DesarrolloValeska Tecas PadillaNo ratings yet
- Mapa Conceptual PDFDocument4 pagesMapa Conceptual PDFmariaNo ratings yet
- Desarrollo Vs MaduracionDocument33 pagesDesarrollo Vs MaduracionJeilin CabreraNo ratings yet
- 5539-Texto Del Artículo-8521-1-10-20090227Document13 pages5539-Texto Del Artículo-8521-1-10-20090227francisco perezNo ratings yet
- Restriccion de Crecimiento Avery 10 EdicionDocument13 pagesRestriccion de Crecimiento Avery 10 EdicionMajito ParraNo ratings yet
- Lectura Ps. Del DesarrolloDocument18 pagesLectura Ps. Del DesarrolloLizzet Magaly Avila ZanabriaNo ratings yet
- Articulo Farmacologia Del Feto y Del Recien Nacido PDFDocument9 pagesArticulo Farmacologia Del Feto y Del Recien Nacido PDFNayely Arrieta GarcíaNo ratings yet
- Determinantes Del Desarrollo y Crecimiento Fetal... JohannDocument1 pageDeterminantes Del Desarrollo y Crecimiento Fetal... JohannGuadalupe Perez PerezNo ratings yet
- Aspectos Grales Del CrecimientoDocument27 pagesAspectos Grales Del Crecimientosusej abigail chaidez zazuetaNo ratings yet
- Crecimiento y Desarrollo Humano 1 (3197)Document92 pagesCrecimiento y Desarrollo Humano 1 (3197)Luz HuamanNo ratings yet
- Expo Rciu LecturaDocument44 pagesExpo Rciu LecturaAna Sofía GómezNo ratings yet
- Capítulos 1 y 3Document49 pagesCapítulos 1 y 3emilio sanchezNo ratings yet
- Cre CimientoDocument40 pagesCre CimientoVale ValeriaNo ratings yet
- Factores Que Influyen en El Crecimiento en La Etapa PrenatalDocument5 pagesFactores Que Influyen en El Crecimiento en La Etapa PrenatalEricson Marchan OlayaNo ratings yet
- Tema O-01 (2002)Document41 pagesTema O-01 (2002)naysha chuquiyauriNo ratings yet
- Mehu525 - U1 - T1 - Características Del CrecimientoDocument22 pagesMehu525 - U1 - T1 - Características Del CrecimientoDiana Carolina ZavaletaNo ratings yet
- Guia 6 Crecimiento y Desarrollo PDFDocument24 pagesGuia 6 Crecimiento y Desarrollo PDFErika Yasmin KrmonaNo ratings yet
- Crecimiento y Desarrollo PuberalDocument14 pagesCrecimiento y Desarrollo PuberalCristina Aracely Muñoz FloresNo ratings yet
- RCIUDocument9 pagesRCIUAnita Gariza SolanoNo ratings yet
- Comparando El Exito Educativo de Corea y FinlandiaDocument9 pagesComparando El Exito Educativo de Corea y FinlandiaJesus David Araujo PalmezanoNo ratings yet
- UD 3. Crecimiento y Desarrollo Infantil Hasta Los 6 AñosDocument23 pagesUD 3. Crecimiento y Desarrollo Infantil Hasta Los 6 Años6p5ztkg52fNo ratings yet
- El Crecimiento Desarrollo y Maduración en La Etapa Prenatal Hasta El Recién NacidoDocument18 pagesEl Crecimiento Desarrollo y Maduración en La Etapa Prenatal Hasta El Recién NacidoRutYT OwONo ratings yet
- Subgrupo 2-PediatríaDocument8 pagesSubgrupo 2-PediatríaMaite PashmayNo ratings yet
- Crecimiento y Desarrollo en PediatriaDocument35 pagesCrecimiento y Desarrollo en PediatriamykarinacaNo ratings yet
- Crecimiento Normal y Patológico Del NiñoDocument15 pagesCrecimiento Normal y Patológico Del NiñoDaniel Vargas AlvaradoNo ratings yet
- TAREA 2 Psicologia Del Desarrollo 1Document16 pagesTAREA 2 Psicologia Del Desarrollo 1Anonymous wAbGYG42D0% (1)
- Factores Que Influyen en El Crecimiento en La Etapa PrenatalDocument7 pagesFactores Que Influyen en El Crecimiento en La Etapa PrenatalGracielaDuarteNo ratings yet
- Crecimiento IntrauterinoDocument5 pagesCrecimiento IntrauterinoFranck PalacinNo ratings yet
- Caracteristicas Del Crecimiento y Desarrollo FisicoDocument16 pagesCaracteristicas Del Crecimiento y Desarrollo FisicoMarco MirandaNo ratings yet
- Consensos - Restriccion Del Crecimiento Intrauterino Perspectiva Endocrinologica 70Document5 pagesConsensos - Restriccion Del Crecimiento Intrauterino Perspectiva Endocrinologica 70VALENTINA CORNELLNo ratings yet
- Crecimiento y Desarrollo Clase 1Document23 pagesCrecimiento y Desarrollo Clase 1gabrielaNo ratings yet
- 04 Fisiología Del CrecimientoDocument12 pages04 Fisiología Del CrecimientoMiguel ArragaNo ratings yet
- Modelo de Boletín-Docente Patricia Zegarra CasasDocument8 pagesModelo de Boletín-Docente Patricia Zegarra CasasAnali Aplaza SalazarNo ratings yet
- Aspectos Nutricionales Gestacion PDFDocument33 pagesAspectos Nutricionales Gestacion PDFRachelNo ratings yet
- Desarrollo PrenatalDocument7 pagesDesarrollo PrenatalEzequiel BeltranNo ratings yet
- Análisis de Un Artículo CientificoDocument3 pagesAnálisis de Un Artículo CientificoJorge CordobaNo ratings yet
- Aspectos Generales de Crecimiento y DesarrolloDocument9 pagesAspectos Generales de Crecimiento y DesarrolloLila Seminario ParedesNo ratings yet
- Presentación Escuela de Arte Infantil Con Garabatos ColoridoDocument38 pagesPresentación Escuela de Arte Infantil Con Garabatos Coloridoesteysi floresNo ratings yet
- APN y Sala de PartoDocument5 pagesAPN y Sala de PartoYasser Calvo GómezNo ratings yet
- Rciu BetDocument50 pagesRciu BetalejarowNo ratings yet
- BIOQUIMICA - ObesidadDocument16 pagesBIOQUIMICA - ObesidadLorena Estefani Cespedes ChavezNo ratings yet
- Obesidad: Información: para PacientesDocument24 pagesObesidad: Información: para PacientesArmando VirtoNo ratings yet
- Osteoartritis Obesidad InflamaciónDocument8 pagesOsteoartritis Obesidad InflamaciónDazamNo ratings yet
- Proyecto de Intervención en Niños y Adolescentes de 10 A 15 Años para La Prevención de La Obesidad A Través de La Plataforma FacebookDocument23 pagesProyecto de Intervención en Niños y Adolescentes de 10 A 15 Años para La Prevención de La Obesidad A Través de La Plataforma FacebookEsteffani RamirezNo ratings yet
- NUTRIGENOMICADocument15 pagesNUTRIGENOMICAJhared Quisbert CuentasNo ratings yet
- Feline Hepatic Lipidosis2Document20 pagesFeline Hepatic Lipidosis2Nicole FuentesNo ratings yet
- Sufre Usted de Resistencia A La LeptinaDocument3 pagesSufre Usted de Resistencia A La LeptinaJorge Alexander Román MoralesNo ratings yet
- Alteración de La Conducta Alimentaria - Intro y EpiDocument17 pagesAlteración de La Conducta Alimentaria - Intro y EpiJuyber Mollinedo QuintoNo ratings yet
- Hormonas Usadas en MedicamentosDocument12 pagesHormonas Usadas en MedicamentosIngrid GarciaNo ratings yet
- El WatDocument19 pagesEl WatAstrid Narro AlvaradoNo ratings yet
- Obesidad ExógenaDocument6 pagesObesidad ExógenaPatricia LópezNo ratings yet
- Preguntas de Nutricion, 1p, 2p y FinalDocument13 pagesPreguntas de Nutricion, 1p, 2p y FinalexaniimaNo ratings yet
- Tesis Factores Laborales Asociados A Sobrepeso y Obesidad en Adultos JóvenesDocument57 pagesTesis Factores Laborales Asociados A Sobrepeso y Obesidad en Adultos JóvenesMontserrat MoctezumaNo ratings yet
- El Cerebro Es GourmetDocument2 pagesEl Cerebro Es GourmetFernandoGutierrezBejarNo ratings yet
- Comedor Nocturno CompletoDocument11 pagesComedor Nocturno CompletoJosselynSánchezYRichardBarrezueta100% (1)
- OBESIDADDocument25 pagesOBESIDADEdgar Manuel Siguas Garcia0% (1)
- Obesidad y SobrepesoDocument96 pagesObesidad y SobrepesoLina HernandezNo ratings yet
- Relacion Del Indice Cintura Cadera PDFDocument69 pagesRelacion Del Indice Cintura Cadera PDFCesar Ortiz VazquezNo ratings yet
- Hipertensión Intracraneal Idiopática (Pseudotumor Cerebri) - Epidemiología y Patogenia - UpToDateDocument19 pagesHipertensión Intracraneal Idiopática (Pseudotumor Cerebri) - Epidemiología y Patogenia - UpToDatefreshaNo ratings yet
- Documento VeterinarioDocument54 pagesDocumento VeterinarioOctavio Bernabe ArevaloNo ratings yet