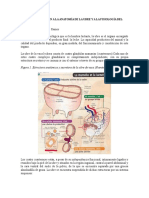
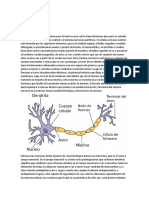
You might also like
- Secrecion de Leche PDFDocument4 pagesSecrecion de Leche PDFMax Lewiss Mafaldo Davila100% (1)
- Anatomia de La Glandula Mamaria 5ADocument30 pagesAnatomia de La Glandula Mamaria 5AAndrea SantosNo ratings yet
- Glandula MamariaDocument12 pagesGlandula MamariaVenezuelaGanadera.comNo ratings yet
- LABORATORIO 3 LecheriaDocument12 pagesLABORATORIO 3 LecheriaGustavo Adolfo PolancoNo ratings yet
- Anatomía de Las Glándulas MamariasDocument40 pagesAnatomía de Las Glándulas MamariasGuillermo Salinas LiendoNo ratings yet
- Anatomía de La Ubre y A La Fisiología Del OrdeñoDocument28 pagesAnatomía de La Ubre y A La Fisiología Del OrdeñoEndrinaYamilethSosaBlancoNo ratings yet
- Ud 4 Anatomia y Fisiologia de La Lactacion PDFDocument45 pagesUd 4 Anatomia y Fisiologia de La Lactacion PDFjosue piquero casoNo ratings yet
- La UbreDocument9 pagesLa UbreAndrea CardozoNo ratings yet
- Caracteristicas Deseables de Una Glandula MamariaDocument36 pagesCaracteristicas Deseables de Una Glandula MamariamariaNo ratings yet
- Fisiología y Anatomía de Las Glándulas Mamarias de La VacaDocument10 pagesFisiología y Anatomía de Las Glándulas Mamarias de La VacaJosue LopezNo ratings yet
- Anatomia y Fisiologia de La Glandula Ma Maria WordDocument6 pagesAnatomia y Fisiologia de La Glandula Ma Maria WordWilderNo ratings yet
- ExamenDocument72 pagesExamenMonykaPaulinaChangoNo ratings yet
- Anatomia y Fisiologia de La Glandula MamDocument9 pagesAnatomia y Fisiologia de La Glandula MamLizeth MenaNo ratings yet
- Secrecion de Leche Por La Ubre de Una Vaca LecheraDocument7 pagesSecrecion de Leche Por La Ubre de Una Vaca LecheraevelinNo ratings yet
- Síntesis y Secreción de LecheDocument4 pagesSíntesis y Secreción de LecheEstefania GodoyNo ratings yet
- Glandulas MamariasDocument56 pagesGlandulas MamariasEfraín TelloNo ratings yet
- Anatomía de La Glándula MamariaDocument5 pagesAnatomía de La Glándula Mamariafabian jara perezNo ratings yet
- Anatomía de La Ubre y A La Fisiología Del OrdeñoDocument3 pagesAnatomía de La Ubre y A La Fisiología Del OrdeñoErick Nava MonterrubioNo ratings yet
- RESUMEN Producción LacteaDocument5 pagesRESUMEN Producción LacteaVale MadrigalNo ratings yet
- Anatomia y Fisiologia de La UbreDocument4 pagesAnatomia y Fisiologia de La UbreSergio TarazonaNo ratings yet
- Trabajo GLANDULA MAMARIADocument19 pagesTrabajo GLANDULA MAMARIAMaria Claudia Pacheco CorchoNo ratings yet
- Anatomía y Fisiología de La Ubre. Calidad Higiénica de La LecheDocument23 pagesAnatomía y Fisiología de La Ubre. Calidad Higiénica de La LecheANANo ratings yet
- Glandula Mamaria 2022Document8 pagesGlandula Mamaria 2022Ninel TorresNo ratings yet
- La Glándula Mamaria Estructura Interna y ExtrernaDocument3 pagesLa Glándula Mamaria Estructura Interna y ExtrernaGenaro Calle ZárateNo ratings yet
- Anatomía de La Glándula MamariaDocument10 pagesAnatomía de La Glándula MamariaAndriyuli Katerin Rebaza ArteagaNo ratings yet
- Glándula MamariaDocument8 pagesGlándula MamariaEDINSON RODRIGO ALVA SEGURANo ratings yet
- Autoevaluación Glándula Mamaria. - Revisión Del IntentoDocument8 pagesAutoevaluación Glándula Mamaria. - Revisión Del IntentoCamilaNo ratings yet
- 18-Conteo CelulasDocument34 pages18-Conteo CelulasSalvadorMartinezToscanoNo ratings yet
- Breve Introducción A La Anatomía de La Ubre y A La Fisiología Del OrdeñoDocument6 pagesBreve Introducción A La Anatomía de La Ubre y A La Fisiología Del OrdeñoBlanca Lorena CampohermosoNo ratings yet
- Estructura de La Glándula MamariaDocument14 pagesEstructura de La Glándula MamariaYAJAIRA EVELIN HOLGUIN GUTIERREZNo ratings yet
- Glandula Mamaria YrDocument9 pagesGlandula Mamaria YrLaura PayaresNo ratings yet
- EMBRIOLOG+ìA, HISTOLOG+ìA Y ANATOM+ìA DE LA MAMADocument9 pagesEMBRIOLOG+ìA, HISTOLOG+ìA Y ANATOM+ìA DE LA MAMAoscarydalcyNo ratings yet
- Glandula MamariaDocument35 pagesGlandula MamariaBayron BenítezNo ratings yet
- Anatomía y Fisiologia de La Glandula MamariaDocument13 pagesAnatomía y Fisiologia de La Glandula MamariaKriss David DelgadoNo ratings yet
- Glándulas de Montgomery Montgomery (Sudoríparas) Por Otro Lado, en El Pezón Existen Muchas TerminacionesDocument7 pagesGlándulas de Montgomery Montgomery (Sudoríparas) Por Otro Lado, en El Pezón Existen Muchas TerminacionesJonathan ValdiviesoNo ratings yet
- Glandulas MamariasDocument8 pagesGlandulas MamariasEstefany SuárezNo ratings yet
- 02-Glándula MamariaDocument12 pages02-Glándula MamariaM RNo ratings yet
- Función de Las Partes de La MamaDocument1 pageFunción de Las Partes de La MamaValeria VarillasNo ratings yet
- Anatomia y Fisiologia de La Glandula MamariaDocument13 pagesAnatomia y Fisiologia de La Glandula MamariaMARYVONNo ratings yet
- 2016nueva Biosintesis de La LecheDocument64 pages2016nueva Biosintesis de La LecheMonse AguilarNo ratings yet
- Fisiologia de La Lactancia MaternaDocument12 pagesFisiologia de La Lactancia MaternaGrace ArevaloNo ratings yet
- Anatomia y Fisiologia de La UbreDocument38 pagesAnatomia y Fisiologia de La UbreAlexDreyfusAnguloNo ratings yet
- Anatomía y Fisiología de La Glándula MamariaDocument5 pagesAnatomía y Fisiología de La Glándula MamariaCarlos Irigoin JaraNo ratings yet
- MAMOGÉNESISDocument14 pagesMAMOGÉNESISalondra martinezNo ratings yet
- Secreción de La Leche y Aspectos BioquimicosDocument28 pagesSecreción de La Leche y Aspectos Bioquimicoslusa fernanda jimenez castellanosNo ratings yet
- Mamas - LactanciaDocument16 pagesMamas - LactanciaNicoli VoltoliniNo ratings yet
- Anatomía de Las MamasDocument8 pagesAnatomía de Las MamasArianna carvajalNo ratings yet
- Glandula MamariaDocument11 pagesGlandula MamariaSofia Reinoso CastilloNo ratings yet
- Biosintesis de La LecheDocument34 pagesBiosintesis de La Lechealexvera69No ratings yet
- Anatomia Aparato Reproductor de La HembraDocument9 pagesAnatomia Aparato Reproductor de La HembraJoss RecaldeNo ratings yet
- 1 Biosintesis y Fase de ProduccionDocument59 pages1 Biosintesis y Fase de ProduccionAnahi RojasNo ratings yet
- Anatomía y Fisiología de La MamaDocument33 pagesAnatomía y Fisiología de La MamaMauRodriguezCardoso100% (2)
- Biosintesis y Fase de ProduccionDocument59 pagesBiosintesis y Fase de Produccionkar KimNo ratings yet
- Glándulas MamariasDocument7 pagesGlándulas MamariasRouz YaiipenNo ratings yet
- (PDF) Anatomia y Fisiologia de La Glándula Mamaria de Animales DomesticosDocument45 pages(PDF) Anatomia y Fisiologia de La Glándula Mamaria de Animales DomesticosAgosto SeptiembreNo ratings yet
- APUNTES - Clínica de BovinosDocument52 pagesAPUNTES - Clínica de BovinosJesús FraustoNo ratings yet
- Anatomia y Fisiologia de La Glandula MamariaDocument7 pagesAnatomia y Fisiologia de La Glandula MamariaCarmen Rosa Ortuño PachecoNo ratings yet
- MF0712_1 - Operaciones auxiliares en reproducción ganaderaFrom EverandMF0712_1 - Operaciones auxiliares en reproducción ganaderaNo ratings yet
- RESUMEN FISIOLOGIA 1 ExamenDocument101 pagesRESUMEN FISIOLOGIA 1 ExamenFavio Alexis DíazNo ratings yet
- Sistema Nervioso 1 ClaseDocument10 pagesSistema Nervioso 1 ClaseEsau GaleanaNo ratings yet
- ACETILACIÓNDocument15 pagesACETILACIÓNJuan Jose Martinez AlcalaNo ratings yet
- Ssss 11111111111Document24 pagesSsss 11111111111Rimberto Quispe AlanocaNo ratings yet
- Bios Bioquim 4 A HamesDocument386 pagesBios Bioquim 4 A HamesPatrick Tejada Adriazola100% (1)
- Taller 3 Celulas Animales y VegetalesDocument16 pagesTaller 3 Celulas Animales y VegetalesRUT HELLENA BATISTA NARVAEZ ESTUDIANTE ACTIVONo ratings yet
- Modulo de Estudio Curso GeneticaDocument150 pagesModulo de Estudio Curso GeneticaJhonson Power Andrade Ordoñez50% (2)
- CitoplasmaDocument11 pagesCitoplasmaManuel AlejandroNo ratings yet
- Actividad Pág. #46 y #47 Biología Aracely Rodríguez PDFDocument7 pagesActividad Pág. #46 y #47 Biología Aracely Rodríguez PDFBryan75% (4)
- La Célula y Sus ComponentesDocument21 pagesLa Célula y Sus Componentesantonia lopezNo ratings yet
- Citoplasma CelularDocument55 pagesCitoplasma CelularVictor Vasquez AvalosNo ratings yet
- Guia de PracticaDocument70 pagesGuia de PracticaEfrain Einer Condori NestasNo ratings yet
- Célula Tipos, Estructura y FuncionesDocument11 pagesCélula Tipos, Estructura y FuncionesNadia GonzálezNo ratings yet
- La Celula HumanaDocument13 pagesLa Celula HumanaGuille Alvarenga100% (1)
- La Celula y Sus OrganelosDocument26 pagesLa Celula y Sus OrganelosOrenLukoNo ratings yet
- Adaptación, Lesion y Muerte CelularDocument32 pagesAdaptación, Lesion y Muerte CelularROSY_LABRONTG100% (1)
- Ap Iiiºmedios Biologia Molecular y Celular - Solucionario Guia Nº1Document4 pagesAp Iiiºmedios Biologia Molecular y Celular - Solucionario Guia Nº1Carmen NeiraNo ratings yet
- Sistema de EndomembranasDocument22 pagesSistema de EndomembranasRubén ChiribogaNo ratings yet
- Texto de Apoyo de Ciencias Naturales 2023Document42 pagesTexto de Apoyo de Ciencias Naturales 2023crknestor849No ratings yet
- La CélulaDocument6 pagesLa CélulaJoseliñha Sanchez GuerraNo ratings yet
- Estructura de La Celula VegetalDocument56 pagesEstructura de La Celula VegetalPepe ZF100% (1)
- Ficha Tarea 4-5 Biología 3°añoDocument3 pagesFicha Tarea 4-5 Biología 3°añocarolinaNo ratings yet
- Guia 5 Actividad Celular PDFDocument10 pagesGuia 5 Actividad Celular PDFChristian BahirNo ratings yet
- Cardiolipidologia PDFDocument198 pagesCardiolipidologia PDFPedro Orlando Saez InostrozaNo ratings yet
- Aparato de Golgi.Document18 pagesAparato de Golgi.Liliana Bolivar GuizarNo ratings yet
- RibosomasDocument4 pagesRibosomasCynthia TijerinaNo ratings yet
- Tema 1 Una Función Química La Vida - 2021Document37 pagesTema 1 Una Función Química La Vida - 2021NataliaNo ratings yet
- Guia Celula CrucigramaDocument8 pagesGuia Celula CrucigramaLady Johanna Conde NietoNo ratings yet
- Sesion de Aprendizaje N°09 Diseño de Prototipo de Celula VegetalDocument6 pagesSesion de Aprendizaje N°09 Diseño de Prototipo de Celula Vegetalwilfredo cajas alvarezNo ratings yet
- Ficha de Reforzamiento 1Document5 pagesFicha de Reforzamiento 1Shirley Pinedo50% (2)