You might also like
- Pathophysiology of SeizuresDocument11 pagesPathophysiology of SeizuresironNo ratings yet
- Question # 1 Identify Patient Problem and Discuss PathophysiologyDocument2 pagesQuestion # 1 Identify Patient Problem and Discuss PathophysiologyzzNo ratings yet
- Pathophysiology of Seizure DisoredersDocument70 pagesPathophysiology of Seizure DisoredersChanchalNo ratings yet
- Intro of Epilepsy - 1. Basic MechanismDocument26 pagesIntro of Epilepsy - 1. Basic MechanismMichael DrakeNo ratings yet
- What Is A SeizureDocument1 pageWhat Is A SeizureSiegfried Punzalan NuñezNo ratings yet
- Pathophysiology of EpilepsyDocument16 pagesPathophysiology of EpilepsyYexalen Zazil100% (1)
- Networks in Temporal Lobe Epilepsy: Karina A. González Otárula,, Stephan SchueleDocument9 pagesNetworks in Temporal Lobe Epilepsy: Karina A. González Otárula,, Stephan SchueleDonald CabreraNo ratings yet
- 2010 Dudek - Primary and Secondary Mechanisms of Epileptogenesis in The Temporal Lobe There Is A Before and An AfterDocument8 pages2010 Dudek - Primary and Secondary Mechanisms of Epileptogenesis in The Temporal Lobe There Is A Before and An AfterfnmendoncaNo ratings yet
- Pathophysiology of Epilepsy: An Updated Review: International Journal of Medical and Health ResearchDocument5 pagesPathophysiology of Epilepsy: An Updated Review: International Journal of Medical and Health ResearchGayathiri GovindarajuNo ratings yet
- Neuro Qu Mic Adela EpilepsiaDocument10 pagesNeuro Qu Mic Adela EpilepsiaKatiuska Yaritza Renteria ValdiviesoNo ratings yet
- Differences: Biological Between Ischemia, Hypoglycemia, EpilepsyDocument9 pagesDifferences: Biological Between Ischemia, Hypoglycemia, EpilepsyAyie JayaNo ratings yet
- Untangling A Web Basic Mechanisms of The Complex Interactions Between Sleep, Circadian Rhythms, and EpilepsyDocument6 pagesUntangling A Web Basic Mechanisms of The Complex Interactions Between Sleep, Circadian Rhythms, and EpilepsyFan AccountNo ratings yet
- Mechanism: OtherDocument2 pagesMechanism: OtherNatia SaginashviliNo ratings yet
- Astrocytes and Epilepsy: Nihal C. de Lanerolle, Tih-Shih Lee, and Dennis D. SpencerDocument15 pagesAstrocytes and Epilepsy: Nihal C. de Lanerolle, Tih-Shih Lee, and Dennis D. SpencerErsin KarataşNo ratings yet
- Learning Issue 1 BAB 2 LaporanDocument32 pagesLearning Issue 1 BAB 2 LaporanHelmiNo ratings yet
- Temporal Lobe Epilepsy - Pathophysiology and Mechanisms: EditorialDocument2 pagesTemporal Lobe Epilepsy - Pathophysiology and Mechanisms: EditorialMahesa Kurnianti PutriNo ratings yet
- The Generation Mechanism of Spike-And-Slow Wave Discharges Appearing On Thalamic Relay NucleiDocument13 pagesThe Generation Mechanism of Spike-And-Slow Wave Discharges Appearing On Thalamic Relay NucleiDaniel TorresNo ratings yet
- Journal of Neurochemistry - 2006 - Bagdy - Serotonin and EpilepsyDocument17 pagesJournal of Neurochemistry - 2006 - Bagdy - Serotonin and EpilepsyFelipe RibeiroNo ratings yet
- Neurometabolic CascadeDocument8 pagesNeurometabolic CascadeTim BonineNo ratings yet
- Ach and SeizureDocument14 pagesAch and Seizurepresentator dadakanNo ratings yet
- Epilepsy AssignmentDocument58 pagesEpilepsy Assignmentnasir khanNo ratings yet
- AS Bases Celulares DE LAS Oscilaciones Neuronales: UmmaryDocument8 pagesAS Bases Celulares DE LAS Oscilaciones Neuronales: Ummarydavid alejandro pinzon sanchezNo ratings yet
- Homeostasis of Intrinsic Excitability in Hippocampal Neurones: Dynamics and Mechanism of The Response To Chronic DepolarizationDocument34 pagesHomeostasis of Intrinsic Excitability in Hippocampal Neurones: Dynamics and Mechanism of The Response To Chronic DepolarizationFedrmNo ratings yet
- Koh - Non-Cell Autonomous Epileptogenesis in Focal Cortical DysplasiaDocument15 pagesKoh - Non-Cell Autonomous Epileptogenesis in Focal Cortical DysplasiakudlaceksystemNo ratings yet
- Epilepsy and Seizures: PathophysiologyDocument11 pagesEpilepsy and Seizures: PathophysiologyAbrizan HassanNo ratings yet
- Ingles, Síndrome ConvulsivoDocument8 pagesIngles, Síndrome ConvulsivoChristian FarinangoNo ratings yet
- Neuro PlasticityDocument67 pagesNeuro Plasticitydrkadiyala2100% (1)
- Leong Tung Ong - Temporal Lobe Epilepsy-Pathophysiology and MechanismsDocument4 pagesLeong Tung Ong - Temporal Lobe Epilepsy-Pathophysiology and MechanismsMarina ShinkoNo ratings yet
- Neuronal Networks in Brain Function, CNS Disorders, and TherapeuticsFrom EverandNeuronal Networks in Brain Function, CNS Disorders, and TherapeuticsCarl FaingoldNo ratings yet
- Jurnal ReadingDocument34 pagesJurnal ReadingFerdian HayafiNo ratings yet
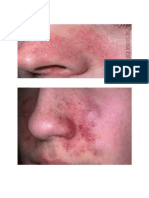
- Facial SeborrheaDocument16 pagesFacial SeborrheaAlejandro ThomasNo ratings yet
- Modulation of Brain Hyperexcitability: Potential New Therapeutic Approaches in Alzheimer's DiseaseDocument37 pagesModulation of Brain Hyperexcitability: Potential New Therapeutic Approaches in Alzheimer's DiseaseVicko SuswidiantoroNo ratings yet
- Epilepsy and Newer Drugs.Document70 pagesEpilepsy and Newer Drugs.Ch Rajesh100% (1)
- Epc 13771Document3 pagesEpc 13771Van Basten ArdiansyachNo ratings yet
- Epilepsy: Ever-Changing States of Cortical Excitability: R. A. B. Badawy, D. R. Freestone, A. Lai AND M. J. CookDocument11 pagesEpilepsy: Ever-Changing States of Cortical Excitability: R. A. B. Badawy, D. R. Freestone, A. Lai AND M. J. CooknurhikmahNo ratings yet
- Computational Models of Neuron-Astrocyte Interaction in EpilepsyDocument10 pagesComputational Models of Neuron-Astrocyte Interaction in EpilepsyEmma FineNo ratings yet
- 32 (Neuroplasticity)Document49 pages32 (Neuroplasticity)SardarChangezKhanNo ratings yet
- Physiology 5 - EpilepsyDocument17 pagesPhysiology 5 - EpilepsyTomgfgd RogestoreNo ratings yet
- Subcortical Epilepsy?: Views & ReviewsDocument7 pagesSubcortical Epilepsy?: Views & ReviewsLuisFer HoundNo ratings yet
- Daniel O. Kellett Et Al - Memory Consolidation in The Cerebellar CortexDocument13 pagesDaniel O. Kellett Et Al - Memory Consolidation in The Cerebellar CortexCortate15gNo ratings yet
- EpilepsyDocument24 pagesEpilepsysalmanhabeebek100% (2)
- 111 Week 2 WorksheetDocument4 pages111 Week 2 WorksheetjabyleynesNo ratings yet
- Epilepsy Principles and DiagnosisDocument26 pagesEpilepsy Principles and DiagnosisLauriettaNo ratings yet
- Gaba Actions and Ionic Plasticity in Epilepsy: SciencedirectDocument8 pagesGaba Actions and Ionic Plasticity in Epilepsy: SciencedirectMuhammad Bayu Zohari HutagalungNo ratings yet
- Se FisiopatoDocument28 pagesSe Fisiopatocska398No ratings yet
- Ni Hms 437509Document19 pagesNi Hms 437509diijah678No ratings yet
- Molecular Pathophysiology of StrokeDocument10 pagesMolecular Pathophysiology of StrokeKahubbi Fatimah Wa'aliyNo ratings yet
- Epilepsy: Head TraumaDocument17 pagesEpilepsy: Head TraumaAimanRiddleNo ratings yet
- Myasthenia Gravis - PathophysiologyDocument3 pagesMyasthenia Gravis - PathophysiologyDill BadaróNo ratings yet
- Gating + HomeostaticDocument7 pagesGating + HomeostaticGhea Mae VillasNo ratings yet
- Overview of Physiology Ayoka AO-1Document38 pagesOverview of Physiology Ayoka AO-1oreoluwa emmanuelNo ratings yet
- Cerri Et Al. 2017Document7 pagesCerri Et Al. 2017JD SánchezNo ratings yet
- Article32 PDFDocument4 pagesArticle32 PDFdupuytrenNo ratings yet
- How Electroconvulsive Therapy Works - Understanding The Neurobiological Mechanisms - PMCDocument23 pagesHow Electroconvulsive Therapy Works - Understanding The Neurobiological Mechanisms - PMCSuraj DubeyNo ratings yet
- Muhlhofer 2018Document15 pagesMuhlhofer 2018bozasnachoNo ratings yet
- Physiology, Neuromuscular Junction of Skeletal MuscleDocument8 pagesPhysiology, Neuromuscular Junction of Skeletal Musclemethmal subasingheNo ratings yet
- 481 FullDocument12 pages481 FullRaaj SharmaNo ratings yet
- Applied Neurophysiology: With Particular Reference to AnaesthesiaFrom EverandApplied Neurophysiology: With Particular Reference to AnaesthesiaNo ratings yet
- From Molecules to Networks: An Introduction to Cellular and Molecular NeuroscienceFrom EverandFrom Molecules to Networks: An Introduction to Cellular and Molecular NeuroscienceRuth HeidelbergerRating: 5 out of 5 stars5/5 (1)
- Electrophysiology Measurements for Studying Neural InterfacesFrom EverandElectrophysiology Measurements for Studying Neural InterfacesNo ratings yet
- Living Mandala TheoryDocument21 pagesLiving Mandala TheoryRandolph Fabian DirectoNo ratings yet
- Enhancement of Heat Exchanger Performance Using Oscillating FlowDocument5 pagesEnhancement of Heat Exchanger Performance Using Oscillating FlowPradyumna DhamangaonkarNo ratings yet
- Secrets of Passion (PDFDrive)Document147 pagesSecrets of Passion (PDFDrive)Hartoyo GondomuliaNo ratings yet
- 11 ENG02 001 (Olamide) PDFDocument62 pages11 ENG02 001 (Olamide) PDFOlamide AdebayoNo ratings yet
- Flow State Brainwave Entrainment GuideDocument36 pagesFlow State Brainwave Entrainment Guidekriskris12345No ratings yet
- Neural DarwinismDocument11 pagesNeural DarwinismRodrigoPristaNo ratings yet
- Us 5935054Document14 pagesUs 5935054danielgoba84No ratings yet
- ElectroencephalographyDocument17 pagesElectroencephalographysambadeeNo ratings yet
- Basics of NeurofeedbackDocument4 pagesBasics of Neurofeedbackapi-372126050% (2)
- Revolutionizing Human-Computer Interaction Through Brain-Machine Interface An EEG-Based ApproachDocument11 pagesRevolutionizing Human-Computer Interaction Through Brain-Machine Interface An EEG-Based ApproachIJRASETPublicationsNo ratings yet
- Thesis Brain Computer InterfaceDocument8 pagesThesis Brain Computer InterfaceCheapCustomWrittenPapersSingapore100% (2)
- Pathophysiology of EpilepsyDocument16 pagesPathophysiology of EpilepsyYexalen Zazil100% (1)
- JC - How Sleep WorksDocument40 pagesJC - How Sleep WorksLe RaNo ratings yet
- The Complementary Nature of Coordination Dynamics-Self-Organization and AgencyDocument8 pagesThe Complementary Nature of Coordination Dynamics-Self-Organization and AgencyNicolás DonosoNo ratings yet
- Module 5 Regulation of Internal EnvironmentDocument17 pagesModule 5 Regulation of Internal EnvironmentSakshi Jauhari100% (1)
- Erol Basar - Memory and Brain Dynamics - Oscillations Integrating Attention, Perception, Learning, and Memory-CRC Press (2004) PDFDocument259 pagesErol Basar - Memory and Brain Dynamics - Oscillations Integrating Attention, Perception, Learning, and Memory-CRC Press (2004) PDFHenry DelgadoNo ratings yet
- Non Invasivebrain ComputerinterfaceforprostheticlimbscontrolinmilitaryDocument14 pagesNon Invasivebrain ComputerinterfaceforprostheticlimbscontrolinmilitaryHayder AdnanNo ratings yet
- Brainwaves and Mind Recent AdvancesDocument90 pagesBrainwaves and Mind Recent AdvancesmkarikanNo ratings yet
- Unlocking The Brain, Volume 1 Coding (Georg Northoff)Document417 pagesUnlocking The Brain, Volume 1 Coding (Georg Northoff)Arbaaz MahmoodNo ratings yet
- 2016 BFsrep38565 MOESM1 ESMDocument14 pages2016 BFsrep38565 MOESM1 ESMAtharv NadkarniNo ratings yet
- A System of Coupled UJT Relaxation Oscil PDFDocument4 pagesA System of Coupled UJT Relaxation Oscil PDFFranwisNo ratings yet
- RitmoDaVidaComoSinfoniaOscilatoria VibracaoSonoraAlteraGeneticaCelularDocument16 pagesRitmoDaVidaComoSinfoniaOscilatoria VibracaoSonoraAlteraGeneticaCelularValdonei SantosNo ratings yet
- Implan Koklea Dan Fungsi Pendengaran (English Version)Document29 pagesImplan Koklea Dan Fungsi Pendengaran (English Version)Julita MelisaNo ratings yet
- Patrick Jemmer - Getting in A (Brain-Wave) State Through Entrainment, Meditation, and HypnosisDocument11 pagesPatrick Jemmer - Getting in A (Brain-Wave) State Through Entrainment, Meditation, and Hypnosistventura100% (1)
- Yoga - Presentation1Document54 pagesYoga - Presentation1ravimars22100% (3)
- System of Binaural BeatsDocument11 pagesSystem of Binaural BeatsGolden SunriseNo ratings yet
- Plagiarism - ReportDocument61 pagesPlagiarism - ReportMahbubul IslamNo ratings yet
- The Complete Guide To Brainwave Entrainment 1Document24 pagesThe Complete Guide To Brainwave Entrainment 1FUMETA100% (2)
- Spontaneous Electrical Activity in The Human Fetal CortexDocument8 pagesSpontaneous Electrical Activity in The Human Fetal CortexFrontiersNo ratings yet
- Nurture Their Nature: Helping Those With A Neurodiversity Enjoy Learning AgainDocument136 pagesNurture Their Nature: Helping Those With A Neurodiversity Enjoy Learning AgainTricia CookNo ratings yet