You might also like
- Ejercicios de Media Mediana y ModaDocument8 pagesEjercicios de Media Mediana y ModaPiro NavarroNo ratings yet
- Formato de Historia ClinicaDocument20 pagesFormato de Historia Clinicaviridianadel100% (1)
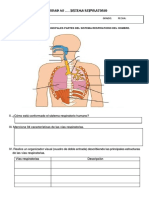
- Ficha de Trabajo Sistema RespiratorioDocument5 pagesFicha de Trabajo Sistema Respiratoriodoris flores colana100% (2)
- Artritis Reumatoide JuvenilDocument7 pagesArtritis Reumatoide JuvenilrxnlrNo ratings yet
- Por Que Los GenogramasDocument4 pagesPor Que Los GenogramasMaye GelvezNo ratings yet
- Seminario de Fisiologia de La GallinaDocument4 pagesSeminario de Fisiologia de La Gallinadany alvarezNo ratings yet
- Medigas FT Aire MedicinalDocument2 pagesMedigas FT Aire MedicinalLuis Centeno UlloaNo ratings yet
- Trauma Ocular y NasalDocument43 pagesTrauma Ocular y NasaljosemerelloNo ratings yet
- Caso Clínico FarmacologiaDocument5 pagesCaso Clínico FarmacologiaGenessis ValdiviezoNo ratings yet
- Tos Con FlemasDocument23 pagesTos Con Flemastito cuadrosNo ratings yet
- Medicamentos VademecumDocument15 pagesMedicamentos VademecumYeison David OrtizNo ratings yet
- Definiciones EPIMIODOLOGIADocument3 pagesDefiniciones EPIMIODOLOGIAcarloschuecos8763No ratings yet
- Direccion de Raices y Conductos Radiculares 2Document18 pagesDireccion de Raices y Conductos Radiculares 2camilo pallesNo ratings yet
- Bioelementos SecundariosDocument5 pagesBioelementos SecundariosLuis Rodriguez AsorzaNo ratings yet
- Formativa #1 Plan de NegociosDocument4 pagesFormativa #1 Plan de NegociosBryan MoralesNo ratings yet
- Asa ProtocoloDocument7 pagesAsa ProtocoloFernanda NavarroNo ratings yet
- Alimentación Complementaria 2018Document28 pagesAlimentación Complementaria 2018Douglas UmbriaNo ratings yet
- El Bovino Criollo en El PeruDocument9 pagesEl Bovino Criollo en El PeruErik Brayan Leyva PérezNo ratings yet
- Ensayo Sobre APSDocument4 pagesEnsayo Sobre APSfabiola salazarNo ratings yet
- Miastenia Gravis: IntroducciónDocument5 pagesMiastenia Gravis: IntroducciónEdgard CubasNo ratings yet
- Medicina InternaDocument1 pageMedicina InternaCarlos HolguinNo ratings yet
- Aspectos Psicológicos Del Hombre en La EnfermedadDocument17 pagesAspectos Psicológicos Del Hombre en La EnfermedadJuan Camilo MedranoNo ratings yet
- 03 Infecciones Urinarias MultiresistenciasDocument24 pages03 Infecciones Urinarias MultiresistenciasRicardo LopezNo ratings yet
- Gestion GebaDocument14 pagesGestion GebaFabian MosqueraNo ratings yet
- Tratamiento para La Enfermedad de ParkinsonDocument20 pagesTratamiento para La Enfermedad de ParkinsonCeci ECNo ratings yet
- Enfermedad de Alzheimer ExposicionDocument21 pagesEnfermedad de Alzheimer ExposicionluisromainNo ratings yet
- AEH SomatizacionDocument23 pagesAEH SomatizacionENRIQUE CAÑADAS RIVERA100% (1)
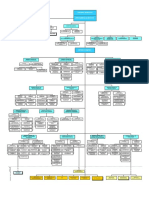
- Organ I GramaDocument1 pageOrgan I GramaAngela Idania Quispe CanorioNo ratings yet
- Uncaria Tomentosa PDFDocument12 pagesUncaria Tomentosa PDFJulie DZNo ratings yet
- El Síncope en PerrosDocument11 pagesEl Síncope en PerrosCarolina Gutiérrez ToroNo ratings yet