You might also like
- DIAGNOSIS Y TAXONOMÍA DE PECESDocument21 pagesDIAGNOSIS Y TAXONOMÍA DE PECESOrtiz PaolaNo ratings yet
- Genero SiphonapteraDocument17 pagesGenero SiphonapteraJenniffer Andrea Gordon EstrellaNo ratings yet
- Conejos y Cuyos (Cunicultura)Document111 pagesConejos y Cuyos (Cunicultura)Vane SolisNo ratings yet
- RotiferaDocument4 pagesRotiferaJorobadoNo ratings yet
- Ser Pien TesDocument31 pagesSer Pien Tescesar romeroNo ratings yet
- III Bim - 5to. Año - Bio - Guía 2 - Reino Animal IIDocument8 pagesIII Bim - 5to. Año - Bio - Guía 2 - Reino Animal IIprofesormiguel241No ratings yet
- SerpentidosDocument37 pagesSerpentidosStefanny Mancero100% (1)
- Actividad 2.2 NSZLDocument13 pagesActividad 2.2 NSZLinfor maticaNo ratings yet
- ZAGREUS HEXACTICTA PPTDocument13 pagesZAGREUS HEXACTICTA PPTAlonzo Aguilar BanegasNo ratings yet
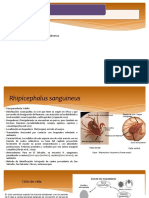
- Rhipicephalus SanguineusDocument4 pagesRhipicephalus SanguineusJeremy RuizNo ratings yet
- Serpientes Yungas 2009 BaldoDocument61 pagesSerpientes Yungas 2009 BaldoMarcelo GutiérrezNo ratings yet
- PRÁCTICA N°9 NEMATODOS ParasitologíaDocument7 pagesPRÁCTICA N°9 NEMATODOS ParasitologíaDavid SilvaNo ratings yet
- Coleópteros forenses: Silphidae y StaphylinidaeDocument32 pagesColeópteros forenses: Silphidae y StaphylinidaeGustavo LimachiNo ratings yet
- Marco Teorico Cuy y HercampuriDocument15 pagesMarco Teorico Cuy y Hercampuricesar eduardoNo ratings yet
- Seminario MoscasDocument30 pagesSeminario MoscasSofía MargaritaNo ratings yet
- Clasificación taxonómica de los insectosDocument110 pagesClasificación taxonómica de los insectosNelson BerriosNo ratings yet
- Tenias en RumiantesDocument49 pagesTenias en RumiantesAlexander Gerardo Calvay Requejo50% (6)
- Ascarididos 0506Document13 pagesAscarididos 0506Jaine LabradaNo ratings yet
- Anfibios: morfología externa y dimorfismo sexualDocument5 pagesAnfibios: morfología externa y dimorfismo sexualmoises balta sandiaNo ratings yet
- Clasificación y características de los peces cartilaginososDocument46 pagesClasificación y características de los peces cartilaginososkatyNo ratings yet
- Introducción Al Cuidado de CobayasDocument26 pagesIntroducción Al Cuidado de Cobayasdavi_ohNo ratings yet
- AnimalesDocument23 pagesAnimalesJ'Kevin Castillo PNo ratings yet
- Ancylostoma y Necator: Características, ciclo y prevención de las uncinariasDocument2 pagesAncylostoma y Necator: Características, ciclo y prevención de las uncinariasKeyla Llontop100% (1)
- Unidad Didactica Vasca PDFDocument26 pagesUnidad Didactica Vasca PDFMiguel A. Fernández MartínezNo ratings yet
- Heterakoideos y Oxyurideos 0506Document6 pagesHeterakoideos y Oxyurideos 0506Giselle Aislinn Lopez MorenoNo ratings yet
- Sesion 4Document25 pagesSesion 4Tiffanny Romero PauccarimaNo ratings yet
- Nematodos AscarisDocument40 pagesNematodos AscarisSegundo Fernandez100% (1)
- PARASITARIASDocument7 pagesPARASITARIASLambari Lizeth Ahuanari GarciaNo ratings yet
- Origen Evolución Caract. Externas.Document57 pagesOrigen Evolución Caract. Externas.alvaro luqueNo ratings yet
- Trabajo Practico N 10Document5 pagesTrabajo Practico N 10Emanuel AlbertoNo ratings yet
- Las BabosasDocument20 pagesLas BabosasCinthya ZambranoNo ratings yet
- Serpiente BotropsDocument41 pagesSerpiente BotropsCrithian HolguinNo ratings yet
- Informe de AnfibiosDocument7 pagesInforme de AnfibioskarenNo ratings yet
- Características de Condrictios RepresentativosDocument6 pagesCaracterísticas de Condrictios RepresentativosOwen Harvey Quispe CasillaNo ratings yet
- Familia OxyuridaeDocument6 pagesFamilia Oxyuridaejavier massellaNo ratings yet
- Tipos de InmadurosDocument12 pagesTipos de InmadurosJefferson SepúlvedaNo ratings yet
- Pulex IrritansDocument7 pagesPulex IrritansSheyla TiconaNo ratings yet
- Orden Diptera 40Document29 pagesOrden Diptera 40Mayra Alexandra Ospina HernandezNo ratings yet
- Gloria Serna 402 EL ESCARABAJODocument20 pagesGloria Serna 402 EL ESCARABAJOglagarbe271No ratings yet
- SerpentesDocument15 pagesSerpentesDaniel Alvarado RobledoNo ratings yet
- Descripcion Biologica de La CodornizDocument8 pagesDescripcion Biologica de La CodornizJorge Chavez100% (1)
- 08 1ESO BG AdaptacionDocument15 pages08 1ESO BG AdaptacionMaria GonzalezNo ratings yet
- Reporte 5 Zoo Henry DimasDocument11 pagesReporte 5 Zoo Henry DimasHenry DLNo ratings yet
- Reproducción tiburones: ovíparos, vivíparos, ovovivíparosDocument2 pagesReproducción tiburones: ovíparos, vivíparos, ovovivíparosNICOLAS DIMETRIC ARIZA PATINONo ratings yet
- Practica 1 de ZoologiaDocument8 pagesPractica 1 de ZoologiaMARIANELLA CORALI LUGO RAMIREZNo ratings yet
- HolothuroideaDocument12 pagesHolothuroideaharryNo ratings yet
- Actividad Semana 5 - ParasitoDocument17 pagesActividad Semana 5 - ParasitoCeciliaNo ratings yet
- Chapulines, Langostas, Grillos y Esperanzas de México Grasshoppers, Locusts, Crickets & Katydids of Mexico Guía Fotográfica Photographic Guide PDFDocument23 pagesChapulines, Langostas, Grillos y Esperanzas de México Grasshoppers, Locusts, Crickets & Katydids of Mexico Guía Fotográfica Photographic Guide PDFLuis RamosNo ratings yet
- 2011 Seleccion Abejas 1Document10 pages2011 Seleccion Abejas 1Mauricio CataniNo ratings yet
- NematodosDocument54 pagesNematodosChrystal Soyoko Chumpitaz Flores100% (1)
- Strep Sip TeraDocument11 pagesStrep Sip TeraALISSON JERALDINNE RODRIGUEZ GIRALDONo ratings yet
- La Evolución de Las SerpientesDocument31 pagesLa Evolución de Las SerpientesVíctor Manuel GonzálezNo ratings yet
- p2.1.b. Generalidades de NematodosDocument11 pagesp2.1.b. Generalidades de NematodosLuis Alfredo Salgado FloresNo ratings yet
- 2do Parcial ParasitoDocument226 pages2do Parcial ParasitoMARIA DE LOS ANGELES CARPIO ARIASNo ratings yet
- Laboratorio # 3ayb Insecta Lepidoptera (Mariposas y Polillas)Document10 pagesLaboratorio # 3ayb Insecta Lepidoptera (Mariposas y Polillas)Geraldinne Trujillo NarvaezNo ratings yet
- Manual de Arañas Universidad de La PlataDocument43 pagesManual de Arañas Universidad de La PlataÓscar Humberto Mejía BlancoNo ratings yet
- TIBURONESDocument14 pagesTIBURONESAllen RmNo ratings yet
- Pags Parasitologia Humana para BioquimicosDocument12 pagesPags Parasitologia Humana para BioquimicosHelena BermeoNo ratings yet
- Nuevo Hoja de Cálculo de Microsoft ExcelDocument1 pageNuevo Hoja de Cálculo de Microsoft ExcelRodrigo VillegasNo ratings yet
- Factor Que Altera El Clima en La SierraDocument1 pageFactor Que Altera El Clima en La SierraRodrigo VillegasNo ratings yet
- Errores Del Metabolismo de CarbohidratosDocument83 pagesErrores Del Metabolismo de CarbohidratosRodrigo VillegasNo ratings yet
- Tesis Marta Fierro 2007Document230 pagesTesis Marta Fierro 2007Rodrigo VillegasNo ratings yet
- MetodologiaDocument9 pagesMetodologiaRodrigo VillegasNo ratings yet
- Botanica Avance 2017Document11 pagesBotanica Avance 2017Rodrigo VillegasNo ratings yet
- Genética Post MendelianaDocument3 pagesGenética Post MendelianaRodrigo VillegasNo ratings yet
- Datos EduDocument4 pagesDatos EduRodrigo VillegasNo ratings yet
- Filo (Phylum) Vs DivisionDocument2 pagesFilo (Phylum) Vs DivisionRodrigo Villegas100% (1)
- MañucoDocument4 pagesMañucoRodrigo VillegasNo ratings yet
- Filo (Phylum) Vs DivisionDocument2 pagesFilo (Phylum) Vs DivisionRodrigo Villegas100% (1)
- Cuantitativa Discreta Grafico de CajasDocument3 pagesCuantitativa Discreta Grafico de CajasRodrigo VillegasNo ratings yet
- 05diagnostico Vol 19 ExtraDocument5 pages05diagnostico Vol 19 ExtraRodrigo VillegasNo ratings yet
- Especificaciones técnicas para construcción de edificioDocument55 pagesEspecificaciones técnicas para construcción de edificioIvonne Del PozoNo ratings yet
- Duwprada - Taller 2Document1 pageDuwprada - Taller 2MARIA ALEJANDRA SAAVEDRANo ratings yet
- Tarea 2Document5 pagesTarea 2Bayron Javier Cortes MartinezNo ratings yet
- Análisis Decreto 579 de 2020Document3 pagesAnálisis Decreto 579 de 2020Sara Castillo VelasquezNo ratings yet
- Manual Thunder Cache Mikrotik CompletoDocument31 pagesManual Thunder Cache Mikrotik CompletoLuis Alberto Gurierrez40% (5)
- Propuesta de Trabajo A DistanciaDocument1 pagePropuesta de Trabajo A DistanciaMaryel LorenzoNo ratings yet
- Cuestionario CompletoDocument6 pagesCuestionario CompletoSarai EspinozaNo ratings yet
- Calentamiento Global Causas EfectosDocument3 pagesCalentamiento Global Causas EfectosDanny montero100% (1)
- ETI - Etapa 1 - Semana 2Document4 pagesETI - Etapa 1 - Semana 2Guillermo AcevedoNo ratings yet
- I - Carga Eléctrica Ley de Coulomb y Principio de Conservacion de La CargaDocument6 pagesI - Carga Eléctrica Ley de Coulomb y Principio de Conservacion de La Cargaanavarro100% (31)
- Acosta-JuanJose - Cuadro Comparativo.Document3 pagesAcosta-JuanJose - Cuadro Comparativo.Maricela CalderonNo ratings yet
- Embargo Retentivo U Opsicion Carlos HerediaDocument6 pagesEmbargo Retentivo U Opsicion Carlos HerediaManuel SantanaNo ratings yet
- AIR Virtual InstrumentsDocument492 pagesAIR Virtual InstrumentsdatttNo ratings yet
- Lab 7. Asesoría, Contratos y Control Disciplinario. Clinica Dto 7 2017Document28 pagesLab 7. Asesoría, Contratos y Control Disciplinario. Clinica Dto 7 2017Roxana Elizabeth López LópezNo ratings yet
- LET-223 Primera Prueba Parcial (1PP-202)Document4 pagesLET-223 Primera Prueba Parcial (1PP-202)joel25% (4)
- Símbolos TuberiasDocument8 pagesSímbolos TuberiasdavidNo ratings yet
- Diseño de buquesDocument53 pagesDiseño de buquesdavalgonzalezNo ratings yet
- Estructura Orgánica de La OMTDocument4 pagesEstructura Orgánica de La OMTJazmín RoblesNo ratings yet
- Webquest CompletoDocument55 pagesWebquest CompletoAldo RomanoNo ratings yet
- Simbolos Electricos Electronicos BasicosDocument5 pagesSimbolos Electricos Electronicos BasicosAntonioNo ratings yet
- Yacimientos Petrolíferos Fiscales BolivianosDocument52 pagesYacimientos Petrolíferos Fiscales Bolivianosdeymar789No ratings yet
- Optimización de producción con Solver en ExcelDocument6 pagesOptimización de producción con Solver en ExcelMartin Castro ArquerosNo ratings yet
- Capilaridad InformeDocument12 pagesCapilaridad InformeHL100% (1)
- Análisis transformador monofásicoDocument6 pagesAnálisis transformador monofásicoJosé IbarraNo ratings yet
- Sociedad del Conocimiento y Contextos Educativos RuralesDocument14 pagesSociedad del Conocimiento y Contextos Educativos RuralesKellys Johana Villarreal Burgos100% (1)
- Reglamentode EnchufesytomacorrientesDocument33 pagesReglamentode EnchufesytomacorrientesSegundo Teofilo Cadenillas CabanillasNo ratings yet
- EESPP Túpac Amaru informe prácticas estudianteDocument2 pagesEESPP Túpac Amaru informe prácticas estudianteJuan Cesar Quispe ArteagaNo ratings yet
- Trabajo Final - Crucigrama Con RespuestasDocument2 pagesTrabajo Final - Crucigrama Con RespuestasTami LiendoNo ratings yet
- CONTRATO DE COMPRAVENTA DE LOTE DE TERRENO - Alvaro AntonioDocument1 pageCONTRATO DE COMPRAVENTA DE LOTE DE TERRENO - Alvaro Antoniopatricia toncelNo ratings yet
- Tarea#1 Compromiso PedagógicoDocument4 pagesTarea#1 Compromiso Pedagógicosara sanchezNo ratings yet