You might also like
- Tarea 4 - ListoDocument3 pagesTarea 4 - ListoAbrekNo ratings yet
- Desarrollo Sustentable en México 1980-2007Document13 pagesDesarrollo Sustentable en México 1980-2007Roberto Moreno JNo ratings yet
- Guía de estudio: Desarrollo sustentableDocument4 pagesGuía de estudio: Desarrollo sustentableAbrekNo ratings yet
- Estrategias para crear valor en la industria de la panificación artesanal peruanaDocument3 pagesEstrategias para crear valor en la industria de la panificación artesanal peruanaAbrek100% (1)
- Tarea 1 AnalisisDocument1 pageTarea 1 AnalisisAbrekNo ratings yet
- Tarea 4 - DecisionesDocument5 pagesTarea 4 - DecisionesAbrekNo ratings yet
- Actividad 5Document6 pagesActividad 5AbrekNo ratings yet
- Estrategias para crear valor en la industria de la panificación artesanal peruanaDocument3 pagesEstrategias para crear valor en la industria de la panificación artesanal peruanaAbrek100% (1)
- Tarea 3 Lenguaje EnsambladorDocument2 pagesTarea 3 Lenguaje EnsambladorAbrek0% (1)
- Tarea 2-ListaDocument7 pagesTarea 2-ListaAbrek100% (1)
- Tarea6 POODocument6 pagesTarea6 POOAbrekNo ratings yet
- Los Facsímiles Figura Por FiguraDocument41 pagesLos Facsímiles Figura Por FiguraRoberto Vinett100% (1)
- Tarea 1Document4 pagesTarea 1Alexander Martinez100% (2)
- Actividad 5Document6 pagesActividad 5AbrekNo ratings yet
- LeyendasDocument3 pagesLeyendasAbrekNo ratings yet
- Receptores DomaminergicosDocument9 pagesReceptores DomaminergicosGuillermo MartinezNo ratings yet
- Los Secretos de Las Platas PDFDocument200 pagesLos Secretos de Las Platas PDFMichelle Nicolett Salas SepulvedaNo ratings yet
- V54n2a12 PDFDocument10 pagesV54n2a12 PDFAbrekNo ratings yet
- Cómo elaborar un programa de trabajo en saludDocument8 pagesCómo elaborar un programa de trabajo en saludAbrekNo ratings yet
- ListeriosisDocument14 pagesListeriosisAbrekNo ratings yet
- Escuela Superior de Medicina Veterinaria y ZootecniaDocument7 pagesEscuela Superior de Medicina Veterinaria y ZootecniaAbrekNo ratings yet
- Ciclo de vida SIDocument43 pagesCiclo de vida SIcarolita71No ratings yet
- Saban AsDocument34 pagesSaban AsJosé Manuel Ramos AlonsoNo ratings yet
- Tarea 1 AnalisisDocument1 pageTarea 1 AnalisisAbrekNo ratings yet
- Educación y Neurociencias.Document14 pagesEducación y Neurociencias.Gabriela GonzálezNo ratings yet
- Biotecnología y Etnomedicina para El Desarrollo HumanoDocument10 pagesBiotecnología y Etnomedicina para El Desarrollo HumanoAbrekNo ratings yet
- 4.la Neurociencia Cognitiva Una Ciencia Base PDFDocument5 pages4.la Neurociencia Cognitiva Una Ciencia Base PDFMario Saldivar100% (1)
- Ibanez Garcia Quesonlasneurociencias PDFDocument105 pagesIbanez Garcia Quesonlasneurociencias PDFAbrekNo ratings yet
- B 032 066 - 7 PDFDocument68 pagesB 032 066 - 7 PDFAbrekNo ratings yet
- Circuitos Basicos de ElectronicaDocument62 pagesCircuitos Basicos de ElectronicaORLANDOGAP100% (2)
- Torque y tensión recomendada para pernos estructurales ASTM A325 y A490Document1 pageTorque y tensión recomendada para pernos estructurales ASTM A325 y A4900 ONo ratings yet
- Matematicas Resueltos (Soluciones) Derivadas 2º Bachillerato Opción B-COUDocument52 pagesMatematicas Resueltos (Soluciones) Derivadas 2º Bachillerato Opción B-COUmanuel_valle_4100% (5)
- Informe 1 EdafoDocument15 pagesInforme 1 EdafoMartha Zevallos75% (12)
- Yacimientos Residuales y de Origen SupergenicoDocument20 pagesYacimientos Residuales y de Origen SupergenicoFranz Diaz BuenoNo ratings yet
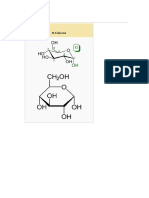
- GlucosaDocument5 pagesGlucosaAdonayDecoracionesNo ratings yet
- Medici OnDocument1 pageMedici OnJesus Manuel MoralesNo ratings yet
- Lab Lodos - DesbloqueadoDocument246 pagesLab Lodos - DesbloqueadoLuis NavarroNo ratings yet
- IND Overview Brochure SpanishDocument35 pagesIND Overview Brochure SpanishJuan Francisco CHavez ViladegutNo ratings yet
- Quimica IDocument242 pagesQuimica ILuis Emanuel Carrera67% (3)
- ElectrostáticaDocument4 pagesElectrostáticaBonitzuZTNo ratings yet
- Informe: Cálculo de Índice de Refracción Del Agua y Distancia Focal de Lentes DelgadasDocument9 pagesInforme: Cálculo de Índice de Refracción Del Agua y Distancia Focal de Lentes DelgadasGaston VilchesNo ratings yet
- Condiciones de Iluminacion en Los Centros de TrabajoDocument15 pagesCondiciones de Iluminacion en Los Centros de TrabajoClaudia BermudezNo ratings yet
- Algebra ExposicionDocument13 pagesAlgebra ExposicionLuizitho BenitezNo ratings yet
- Conceptos básicos de electricidad, corrientes y semiconductoresDocument21 pagesConceptos básicos de electricidad, corrientes y semiconductoresnetpedrocr2070No ratings yet
- Fórmulas básicas del movimiento circularDocument2 pagesFórmulas básicas del movimiento circularEric ArrietaNo ratings yet
- TEODOLITO Y MEDICIÓN DE ÁNGULOSDocument64 pagesTEODOLITO Y MEDICIÓN DE ÁNGULOStifani santosNo ratings yet
- Matrices en ExcelDocument20 pagesMatrices en ExcelFrancis5051No ratings yet
- Clase N°05Document15 pagesClase N°05Lucerito Madeleine Flores ValladaresNo ratings yet
- Planif 1aDocument6 pagesPlanif 1aM Eugenia QuirogaNo ratings yet
- Ejercicios de trabajo y energíaDocument4 pagesEjercicios de trabajo y energíajuniorNo ratings yet
- Trabajo Electrotecnia I-2016 Nuevo Tema 1Document17 pagesTrabajo Electrotecnia I-2016 Nuevo Tema 1Jefferson Leonel Cesar JuarezNo ratings yet
- Tarea 5Document5 pagesTarea 5Shalito MHNo ratings yet
- MdsDocument8 pagesMdsGina Gisella GonzalezNo ratings yet
- Pines Intradentinarios o Espigas PDFDocument12 pagesPines Intradentinarios o Espigas PDFgerardo100% (1)
- Guía de Laboratorio MRUV Física 10°Document1 pageGuía de Laboratorio MRUV Física 10°xitzSkyBluex YT100% (1)
- Informe 2 - 2012 Pa Pegar REAL FINALLDocument9 pagesInforme 2 - 2012 Pa Pegar REAL FINALLDiego Roa CossioNo ratings yet
- Defectos FCAWDocument54 pagesDefectos FCAWWilmer Ramírez Merejildo100% (3)
- Bauhaus Espana Catalogo para El Reformista PDFDocument408 pagesBauhaus Espana Catalogo para El Reformista PDFpelabolas.comVenezuelaNo ratings yet
- Paredes Trabajo IndiviDocument14 pagesParedes Trabajo IndiviAlexMiguelNo ratings yet
- Conservación de cantidad de movimiento ecuaciones líneal y angularDocument64 pagesConservación de cantidad de movimiento ecuaciones líneal y angularAlexander GarcíaNo ratings yet