You might also like
- Mikrobiologi Medis I: Patogen dan Mikrobioma ManusiaFrom EverandMikrobiologi Medis I: Patogen dan Mikrobioma ManusiaRating: 4 out of 5 stars4/5 (11)
- Tugas Paper Oksidasi (Ni Kadek Tini Malini - 21820070)Document11 pagesTugas Paper Oksidasi (Ni Kadek Tini Malini - 21820070)Dexa RamadhaniNo ratings yet
- Mikrobiologi Perubatan I: Patogen dan Mikrobiologi ManusiaFrom EverandMikrobiologi Perubatan I: Patogen dan Mikrobiologi ManusiaRating: 2.5 out of 5 stars2.5/5 (2)
- Probiotik EditDocument16 pagesProbiotik EditAidil PutraNo ratings yet
- Teknik Asas Pembiakan Hibrid Ikan Keli Thailand (Clarias sp.)From EverandTeknik Asas Pembiakan Hibrid Ikan Keli Thailand (Clarias sp.)Rating: 5 out of 5 stars5/5 (2)
- Oli Go Sakari DaDocument25 pagesOli Go Sakari Daakreditasi rscNo ratings yet
- UNTUK METABOLIT SEKUNDER MIKROBADocument17 pagesUNTUK METABOLIT SEKUNDER MIKROBAIfungNo ratings yet
- Laporan Praktikum AntioksidanDocument32 pagesLaporan Praktikum AntioksidanFaranita Lutfia Normasari50% (2)
- UJI AKTIVITAS ENZIMATIK MIKROBADocument4 pagesUJI AKTIVITAS ENZIMATIK MIKROBAViaNo ratings yet
- Laporan Asam LaktatDocument19 pagesLaporan Asam Laktatfun-funNo ratings yet
- Nomor 1 Dan 3BDocument4 pagesNomor 1 Dan 3BYuu WidaaNo ratings yet
- EFEK ANTIOKSIDAN C-FIKOSIANINDocument8 pagesEFEK ANTIOKSIDAN C-FIKOSIANINAldiNo ratings yet
- Laporan Oksidasi BiologiDocument12 pagesLaporan Oksidasi BiologiWahyu BektiNo ratings yet
- Dasar Teori Bakteri Asam LaktatDocument20 pagesDasar Teori Bakteri Asam Laktatrezkinugroho100% (1)
- Aktivitas Antioksidan Tumbuhan Terpilih Dari Ramuan Empiris Untuk Penyakit Liver - Kajian Fisika-Kimia Dan Biologi MolekularDocument4 pagesAktivitas Antioksidan Tumbuhan Terpilih Dari Ramuan Empiris Untuk Penyakit Liver - Kajian Fisika-Kimia Dan Biologi MolekularIsmedsyah SyahNo ratings yet
- TUGAS BIOKIMIA 1 - C - 21820086 - Dexa Ramadhani.Document10 pagesTUGAS BIOKIMIA 1 - C - 21820086 - Dexa Ramadhani.Dexa RamadhaniNo ratings yet
- EKSTRAK TOMAT MENURUNKAN GSHDocument10 pagesEKSTRAK TOMAT MENURUNKAN GSHNury Fitria PiaNo ratings yet
- A - P21118047 - Deice Tresia-1Document6 pagesA - P21118047 - Deice Tresia-1Deice TresyaNo ratings yet
- Aloksan 120Document12 pagesAloksan 120Devi Dwi PanduNo ratings yet
- BAB II Tinjauan PustakaDocument31 pagesBAB II Tinjauan PustakaYondri Mandaku Tasidjawa100% (1)
- GLUKONEOGENESISDocument13 pagesGLUKONEOGENESISnadya paramitaNo ratings yet
- Antioksidan Ujian Tengah SemesterDocument5 pagesAntioksidan Ujian Tengah Semesteraflaham majidNo ratings yet
- Jahe AntioksidanDocument13 pagesJahe AntioksidanPutri Laili AdzaniNo ratings yet
- Biokimia M. Ananta FirdausDocument7 pagesBiokimia M. Ananta FirdausAnanta FirdausNo ratings yet
- Respon ImunDocument6 pagesRespon ImunJoan destalino destalinoNo ratings yet
- Mekanisme Flavonoid Sebagai AntibakteriDocument4 pagesMekanisme Flavonoid Sebagai AntibakteriAndyNo ratings yet
- BAKTERIOSINDocument13 pagesBAKTERIOSINMeyraaniiNo ratings yet
- Aktivitas Fisik Berat Mengakibatkan Degenerasi Berat Sel HepatositDocument5 pagesAktivitas Fisik Berat Mengakibatkan Degenerasi Berat Sel HepatositTiara Grhanesia DenashuryaNo ratings yet
- Adoc - Pub - II Tinjauan Pustaka Ternak Unggas Merupakan JenisDocument10 pagesAdoc - Pub - II Tinjauan Pustaka Ternak Unggas Merupakan JenisU EnNo ratings yet
- G70122097 - Metabolisme Dan Fisiologi MikroorganismeDocument5 pagesG70122097 - Metabolisme Dan Fisiologi MikroorganismeZulfadliNo ratings yet
- S09092fk Perbandingan Aktivitas LiteraturDocument14 pagesS09092fk Perbandingan Aktivitas LiteraturIlham HusniNo ratings yet
- KimpesDocument6 pagesKimpesMahfudz IhsanNo ratings yet
- Oksidasi BiologisDocument6 pagesOksidasi BiologisLuqyana BardaNo ratings yet
- Manajemen Cairan Pasien AnakDocument8 pagesManajemen Cairan Pasien AnakDinar ChiekoNo ratings yet
- ZincDocument3 pagesZincGallant devaNo ratings yet
- ENZIM KATALASEDocument24 pagesENZIM KATALASEYolanda SPNo ratings yet
- Resume UAS NUTRADocument10 pagesResume UAS NUTRASukmamunawwarahNo ratings yet
- 016 - Antioksidan Dan Radikal Bebas - Rahma Dwi ArdianiDocument8 pages016 - Antioksidan Dan Radikal Bebas - Rahma Dwi ArdianiRahma dwiNo ratings yet
- Antioksidan Talas Ungu IVENDocument22 pagesAntioksidan Talas Ungu IVENIvhen AninNo ratings yet
- PROBIOTIK MENYEHATKANDocument6 pagesPROBIOTIK MENYEHATKANReiZitosz Be'elNo ratings yet
- Acetaminophen (Paracetamol) Induced HepatotoxicDocument22 pagesAcetaminophen (Paracetamol) Induced HepatotoxicUsi Wulandari IINo ratings yet
- L. rhamnosus SKG 34 potensi probiotikDocument4 pagesL. rhamnosus SKG 34 potensi probiotikYumailita SetiawatiNo ratings yet
- Efek Rutin Flavonol Pada Kadar Besi Serum Dan HatiDocument10 pagesEfek Rutin Flavonol Pada Kadar Besi Serum Dan Hatiyogi gestiawanNo ratings yet
- OPTIMASI PENGIRADIASIANDocument6 pagesOPTIMASI PENGIRADIASIANRissal RisyandiNo ratings yet
- Laporan Biokimia Enzim KatalaseDocument20 pagesLaporan Biokimia Enzim Katalasehalimatus istiqomahNo ratings yet
- ENZIM DAN KINETIKADocument31 pagesENZIM DAN KINETIKAbiasa agusNo ratings yet
- 385-Article Text-701-1-10-20170414Document5 pages385-Article Text-701-1-10-20170414Evan WalkerNo ratings yet
- Laporan Praktikum Biokimia KatalaseDocument30 pagesLaporan Praktikum Biokimia Katalase노베라No ratings yet
- LEPTIN DAN PERAN IMUNOMETABOLISMEDocument32 pagesLEPTIN DAN PERAN IMUNOMETABOLISMEelsa auliaNo ratings yet
- Bakteri LactobasiusDocument22 pagesBakteri LactobasiusA.M IQBALNo ratings yet
- JUDULDocument8 pagesJUDULNanaNo ratings yet
- ROS role in brucein D-mediated p38 MAPK and NF-kB pathways in pancreatic cancerDocument27 pagesROS role in brucein D-mediated p38 MAPK and NF-kB pathways in pancreatic cancerReenchezya AuliaNo ratings yet
- Vitamin C dan Status Autofagi pada Drosophila melanogaster yang Terinduksi FeSO4Document20 pagesVitamin C dan Status Autofagi pada Drosophila melanogaster yang Terinduksi FeSO4LisaNo ratings yet
- OPTIMALKAN VEGF DALAM HIPERTENSI KEHAMILANDocument10 pagesOPTIMALKAN VEGF DALAM HIPERTENSI KEHAMILANRaniNo ratings yet
- ISONIAZIDDocument8 pagesISONIAZIDPaulus AnungNo ratings yet
- AsamLaktatDocument3 pagesAsamLaktatTassy PutriNo ratings yet
- Garuda 1091372Document5 pagesGaruda 1091372Danang RaharjoNo ratings yet
- FAKTOR PERTUMBUHAN BAKTERIDocument11 pagesFAKTOR PERTUMBUHAN BAKTERIAlhex IntelNo ratings yet
- ANTIOKSIDANDocument28 pagesANTIOKSIDANUmiyati GustikaNo ratings yet
- Presentasi - Hubungan HBOT Dengan ROSDocument32 pagesPresentasi - Hubungan HBOT Dengan ROSRusda SyawieNo ratings yet
- Ulkus KorneaDocument12 pagesUlkus KorneaDoni Andika PutraNo ratings yet
- Anatomi Traktus UrinariusDocument49 pagesAnatomi Traktus UrinariusDoni Andika PutraNo ratings yet
- Jurnal Neuro Brader HafizDocument18 pagesJurnal Neuro Brader HafizDoni Andika PutraNo ratings yet
- Perawatan Nifas Luka CaesarDocument56 pagesPerawatan Nifas Luka CaesarDoni Andika PutraNo ratings yet
- Warna Warni Internsip Di Bungo CityDocument3 pagesWarna Warni Internsip Di Bungo CityDoni Andika PutraNo ratings yet
- Padang Crs Perawatan Luka NifasDocument49 pagesPadang Crs Perawatan Luka NifasDoni Andika PutraNo ratings yet
- Bakti Universitas UsaiDocument1 pageBakti Universitas UsaiRey M FakhruryNo ratings yet
- Hipertensive Heart DiseaseDocument25 pagesHipertensive Heart DiseaseDoni Andika Putra100% (1)
- Daftar Wahana Angkatan I 2020Document5 pagesDaftar Wahana Angkatan I 2020Kevin WewengkangNo ratings yet
- Serba 7 Di UNAND Part 1Document2 pagesSerba 7 Di UNAND Part 1Doni Andika PutraNo ratings yet
- Case Report SessionDocument21 pagesCase Report SessiondilaladyanaNo ratings yet
- Ario Fix Serba 7 Di UNAND Part 2Document4 pagesArio Fix Serba 7 Di UNAND Part 2Doni Andika PutraNo ratings yet
- USU KarsinomaDocument11 pagesUSU KarsinomaFahNo ratings yet
- Case Dra PJR REZADocument19 pagesCase Dra PJR REZADoni Andika PutraNo ratings yet
- Tatalaksana Penyalahgunaan OpioidDocument7 pagesTatalaksana Penyalahgunaan OpioidDoni Andika PutraNo ratings yet
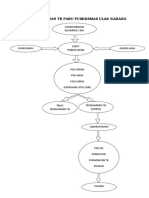
- Alur Pelayanan TB Paru Puskesmas Ulak KarangDocument1 pageAlur Pelayanan TB Paru Puskesmas Ulak KarangDoni Andika PutraNo ratings yet
- Jurnal Epidemiology PJKDocument8 pagesJurnal Epidemiology PJKDoni Andika PutraNo ratings yet
- Kanker Mulut 3Document5 pagesKanker Mulut 3Doni Andika PutraNo ratings yet
- Intrakranial Space Occupying LessionDocument20 pagesIntrakranial Space Occupying LessionDoni Andika PutraNo ratings yet
- Petunjuk Mengaktifkan AlatDocument1 pagePetunjuk Mengaktifkan AlatDoni Andika PutraNo ratings yet
- MENINGITISDocument31 pagesMENINGITISDoni Andika PutraNo ratings yet
- Anemia IdDocument9 pagesAnemia IdDoni Andika PutraNo ratings yet
- MENINGITISDocument31 pagesMENINGITISDoni Andika PutraNo ratings yet
- CRS VerukaDocument21 pagesCRS VerukaDoni Andika PutraNo ratings yet
- 2 1 6Document8 pages2 1 6Aprilia PuspitasariNo ratings yet
- OmskDocument20 pagesOmskYuliana Marbun0% (1)
- Case GnapsDocument38 pagesCase GnapsDoni Andika PutraNo ratings yet
- Hemofilia 1Document26 pagesHemofilia 1Rini Anggraini Hefni PNo ratings yet
- Referat Degenerative Spine Disease-1Document28 pagesReferat Degenerative Spine Disease-1Doni Andika Putra100% (1)