You might also like
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (265)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (587)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (894)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Anesthesia Secret PDFDocument467 pagesAnesthesia Secret PDFPutin Phy100% (8)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- Nursing Process in Drug TherapyDocument60 pagesNursing Process in Drug TherapyYra JhaneNo ratings yet
- USMLE High YieldDocument8 pagesUSMLE High YieldNajia ChoudhuryNo ratings yet
- Clinical Trial in IndiaDocument9 pagesClinical Trial in IndiaBhupesh KumarNo ratings yet
- Russian and US Mind Control Technology Psychotronic WarDocument123 pagesRussian and US Mind Control Technology Psychotronic WardropsyNo ratings yet
- TRS 957 (2010) - Annex 3 - WHO GMP For Pharmaceutical Products Containing Hazardous SubstancesDocument17 pagesTRS 957 (2010) - Annex 3 - WHO GMP For Pharmaceutical Products Containing Hazardous SubstancesQuang Hiếu NgôNo ratings yet
- Understanding Epileptic SeizuresDocument19 pagesUnderstanding Epileptic SeizuresRiki AntoNo ratings yet
- People Pleasing Patterns Are Learned When Needs Are Not Met PDFDocument10 pagesPeople Pleasing Patterns Are Learned When Needs Are Not Met PDFPamela RodriguezNo ratings yet
- Surya Namaskar BenefitsDocument16 pagesSurya Namaskar BenefitsMillion Dollar KnowledgeNo ratings yet
- Remember: Blood CirculationDocument4 pagesRemember: Blood CirculationvicpardoNo ratings yet
- Chest X-Rays:: Case Studies From The FieldDocument12 pagesChest X-Rays:: Case Studies From The FieldRiki AntoNo ratings yet
- ILAE Clasiffication and Teminology 2017Document10 pagesILAE Clasiffication and Teminology 2017Prabaningrum DwidjoasmoroNo ratings yet
- Ankylosing SpondylitesDocument16 pagesAnkylosing SpondylitesJyotirmayeeNo ratings yet
- 113 JMSCRDocument12 pages113 JMSCRRiki AntoNo ratings yet
- Valproic Acid and Epilepsy: From Molecular Mechanisms To Clinical EvidencesDocument21 pagesValproic Acid and Epilepsy: From Molecular Mechanisms To Clinical EvidencesRiki AntoNo ratings yet
- Suspected Small-Bowel ObstructionDocument12 pagesSuspected Small-Bowel ObstructionRiki AntoNo ratings yet
- Cranial Nerves OverviewDocument17 pagesCranial Nerves OverviewRiki AntoNo ratings yet
- MainDocument4 pagesMainRiki AntoNo ratings yet
- Obat Epilepsi PDFDocument24 pagesObat Epilepsi PDFYulia Azkia RahmandaNo ratings yet
- Guidelines Allergic RhinitisDocument29 pagesGuidelines Allergic RhinitisAnnisa Wilda SafinaNo ratings yet
- A Review of Rabies Disease, Its Transmission and TreatmentDocument6 pagesA Review of Rabies Disease, Its Transmission and TreatmentRiki AntoNo ratings yet
- COVID-19 Susceptibility in Bronchial Asthma: Original ArticleDocument10 pagesCOVID-19 Susceptibility in Bronchial Asthma: Original ArticleRiki AntoNo ratings yet
- Fulfillment of The Criteria For Diagnosis of Cephalo-Pelvic Disproportion - ACOG GuidelinesDocument5 pagesFulfillment of The Criteria For Diagnosis of Cephalo-Pelvic Disproportion - ACOG GuidelinesRiki AntoNo ratings yet
- 77-Article Text-176-1-10-20191002Document10 pages77-Article Text-176-1-10-20191002Riki AntoNo ratings yet
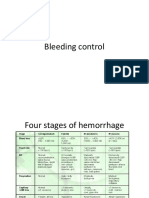
- 4 Bleeding ControlDocument14 pages4 Bleeding ControlRiki AntoNo ratings yet
- Evaluation of Ascites and Its Etiology Using Ultrasonography JRD 119 PDFDocument4 pagesEvaluation of Ascites and Its Etiology Using Ultrasonography JRD 119 PDFEstu Paramadina PratamaNo ratings yet
- The Role of Vitamin A To The EyeDocument7 pagesThe Role of Vitamin A To The EyeRiki AntoNo ratings yet
- Daftar Pustaka Kulit dan KelaminDocument1 pageDaftar Pustaka Kulit dan KelaminRiki AntoNo ratings yet
- S Sssssssssssssssssssss S S: Ipad AirDocument2 pagesS Sssssssssssssssssssss S S: Ipad AirRiki AntoNo ratings yet
- Tugas 2 (Paparan Kimia)Document9 pagesTugas 2 (Paparan Kimia)Nur AnnisaNo ratings yet
- Hib (Who)Document3 pagesHib (Who)Riki AntoNo ratings yet
- 01 GDL Dewiyulino 956 1 Ktidewi 0Document9 pages01 GDL Dewiyulino 956 1 Ktidewi 0Riki AntoNo ratings yet
- Hib (Who)Document3 pagesHib (Who)Riki AntoNo ratings yet
- Referral DiagramDocument1 pageReferral DiagramRiki AntoNo ratings yet
- Dyspnoea: Pathophysiology and A Clinical Approach: ArticleDocument5 pagesDyspnoea: Pathophysiology and A Clinical Approach: ArticleadninhanafiNo ratings yet
- A Vaccine For MalariaDocument2 pagesA Vaccine For MalariaRiki AntoNo ratings yet
- A New Era in Secondary Prevention After Acute Coronary SyndromeDocument3 pagesA New Era in Secondary Prevention After Acute Coronary SyndromeRiki AntoNo ratings yet
- YanBoy Trading Company Profile 2020Document6 pagesYanBoy Trading Company Profile 2020Andile NtuliNo ratings yet
- Fire Retardant Research PaperDocument2 pagesFire Retardant Research Paperapi-318759920No ratings yet
- Model Testimi Në Lëndën Gjuha Angleze, Klasa e 9, Model 1 PDFDocument6 pagesModel Testimi Në Lëndën Gjuha Angleze, Klasa e 9, Model 1 PDFndojafernandagmail.comNo ratings yet
- The Lord of Lost Heart PDFDocument44 pagesThe Lord of Lost Heart PDFCouteron LaurentNo ratings yet
- Analytical Exposition Text Corona Should Be Against Surround UsDocument2 pagesAnalytical Exposition Text Corona Should Be Against Surround UsRifkyNo ratings yet
- Introduction of PhobiaDocument2 pagesIntroduction of PhobiawoodenskyNo ratings yet
- APSACS Summer Holiday Homework Guide Warm Region 2023 24 Dated 02 June 2023 - CompressedDocument17 pagesAPSACS Summer Holiday Homework Guide Warm Region 2023 24 Dated 02 June 2023 - CompressedGaming 7HawkNo ratings yet
- Introduction To Environmental Health LectureDocument41 pagesIntroduction To Environmental Health LectureJudy OuNo ratings yet
- ImplantsDocument11 pagesImplantsraheelNo ratings yet
- Report - AYESHA KHATOON (230750060006)Document3 pagesReport - AYESHA KHATOON (230750060006)ahmedkhandba1No ratings yet
- THHV- 18 G10 ĐỀ NÂNG CAO TỔNG HỢP SỐ 2Document12 pagesTHHV- 18 G10 ĐỀ NÂNG CAO TỔNG HỢP SỐ 2hCby 28No ratings yet
- Macronutrients ProteinsDocument2 pagesMacronutrients ProteinsEllice O. MonizNo ratings yet
- MKWD Lwua Adb Package 2 AnnexesDocument199 pagesMKWD Lwua Adb Package 2 AnnexesRoland AnaumNo ratings yet
- 1 s2.0 S2772569323000026 MainDocument8 pages1 s2.0 S2772569323000026 MainAditya RizkyNo ratings yet
- New Consensus On Non Cirrhotic Portal FibrosisDocument41 pagesNew Consensus On Non Cirrhotic Portal Fibrosisramesh5889No ratings yet
- Grade 7 (TLE) Week 1Document12 pagesGrade 7 (TLE) Week 1RoswlleNo ratings yet
- Buku Koding 1Document74 pagesBuku Koding 1indra_igrNo ratings yet
- Workshop Statistical Methods in Biomedical Research - Nioh - NimsDocument3 pagesWorkshop Statistical Methods in Biomedical Research - Nioh - NimsSudheerChauhanNo ratings yet
- Unit 4 BacteriologyDocument51 pagesUnit 4 BacteriologySharmila LamisharNo ratings yet
- Commissioning Federal Facilities Chiller (PQ)Document19 pagesCommissioning Federal Facilities Chiller (PQ)Yu-Chih PuNo ratings yet
- 22.04.2019 - Confined Space A Dangerous SpaceDocument1 page22.04.2019 - Confined Space A Dangerous SpaceVaibhav Vithoba NaikNo ratings yet
- Kindergarten q1 Week10 v2Document33 pagesKindergarten q1 Week10 v2Aryan Angela Dela CruzNo ratings yet