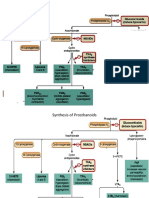
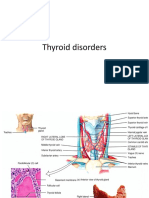
You might also like
- Liver CirrhosisDocument31 pagesLiver CirrhosisMarisol Jane Jomaya50% (2)
- Homeopathic Remedy Pictures Vicki Mathison Frans Kusse.11789 - 1 PDFDocument19 pagesHomeopathic Remedy Pictures Vicki Mathison Frans Kusse.11789 - 1 PDFSrinivas Srinu50% (2)
- Case Study 17Document5 pagesCase Study 17api-27655178367% (3)
- Abdomen Practice Exam - 1Document11 pagesAbdomen Practice Exam - 1Obinna Adolalom100% (1)
- Transplant HepaticDocument409 pagesTransplant HepaticFlorin Calin LungNo ratings yet
- Hepatic Computed Tomography and Cholangiography by Use of Gadoxetic Acid in Healthy CatsDocument11 pagesHepatic Computed Tomography and Cholangiography by Use of Gadoxetic Acid in Healthy CatsAlejandro Estrada RiosNo ratings yet
- Patel 2020Document31 pagesPatel 2020larissa brandaliseNo ratings yet
- Bmri2018 4706976Document11 pagesBmri2018 4706976Denny BunNo ratings yet
- Brown 2000Document12 pagesBrown 2000christian.sincheNo ratings yet
- Reply: Letters To The EditorDocument2 pagesReply: Letters To The Editorgevowo3277No ratings yet
- Jurnal KomparatifDocument9 pagesJurnal KomparatifAulin putri damayantiNo ratings yet
- Excess Parathyroid Hormone Adversely Affects Lipid Metabolism in Chronic Renal FailureDocument5 pagesExcess Parathyroid Hormone Adversely Affects Lipid Metabolism in Chronic Renal FailureJuni ClaudiaNo ratings yet
- Brown 1998Document9 pagesBrown 1998christian.sincheNo ratings yet
- ClinTransplant 2009 DominoLTDocument6 pagesClinTransplant 2009 DominoLTflorinscribdNo ratings yet
- البحث السادسDocument11 pagesالبحث السادسsoha ahmedNo ratings yet
- Art. DanazolDocument9 pagesArt. DanazolmrintraNo ratings yet
- Human Hepatocyte Growth Factor in Blood of Patients With Fulminant Hepatic FailureDocument6 pagesHuman Hepatocyte Growth Factor in Blood of Patients With Fulminant Hepatic FailurePercy SolisNo ratings yet
- Selzner, 2016Document8 pagesSelzner, 2016patriciamendes.contatoNo ratings yet
- p450 LiquinyangDocument4 pagesp450 LiquinyangThoriqotil Haqqul MauludiyahNo ratings yet
- Verboven Moya Gastro 2020Document31 pagesVerboven Moya Gastro 2020Stev Barros ChiribogaNo ratings yet
- Xu 2010 Carbohydrate-To-Fat Ratio Affects Food Intake and Body Weight in Wistar RatsDocument6 pagesXu 2010 Carbohydrate-To-Fat Ratio Affects Food Intake and Body Weight in Wistar RatsPaul SimononNo ratings yet
- Seminario 1A Secrecion BiliarDocument9 pagesSeminario 1A Secrecion BiliarMistic TarotNo ratings yet
- B3 Niacin Energy MitochondriaDocument15 pagesB3 Niacin Energy MitochondriavalgurNo ratings yet
- 17tohj (Ervandi)Document4 pages17tohj (Ervandi)Ervandi AbudiNo ratings yet
- Guillouzo Et Al. - 2007 - The Human Hepatoma HepaRG Cells A Highly Differentiated Model For Studies of Liver Metabolism and Toxicity ofDocument8 pagesGuillouzo Et Al. - 2007 - The Human Hepatoma HepaRG Cells A Highly Differentiated Model For Studies of Liver Metabolism and Toxicity ofyannick brunatoNo ratings yet
- Art 1Document18 pagesArt 1Luis Hurtado PardoNo ratings yet
- Artigo Biotecnologia Tecido AdiposoDocument15 pagesArtigo Biotecnologia Tecido AdiposoLuiz Gabriel Ferreira da SilvaNo ratings yet
- Glutamine Donor Pretreatment in Rat Kidney Transplants With Severe Preservation Reperfusion InjuryDocument7 pagesGlutamine Donor Pretreatment in Rat Kidney Transplants With Severe Preservation Reperfusion InjuryAncaVerdesNo ratings yet
- Hypothermic Machine Preservation in Human Liver Transplantation: The First Clinical SeriesDocument10 pagesHypothermic Machine Preservation in Human Liver Transplantation: The First Clinical SeriesRodolpho ViégasNo ratings yet
- 17TOHJDocument4 pages17TOHJAriffa DiazNo ratings yet
- Etm 14 02 1496Document9 pagesEtm 14 02 1496Bella La Rosa HusnaNo ratings yet
- Research Article: Oxaliplatin But Not Irinotecan Impairs Posthepatectomy Liver Regeneration in A Murine ModelDocument6 pagesResearch Article: Oxaliplatin But Not Irinotecan Impairs Posthepatectomy Liver Regeneration in A Murine ModelDaniel Kings SanyaNo ratings yet
- Ankaflavin Ameliorates Steatotic Liver Ischemia - Reperfusion Injury in MicDocument7 pagesAnkaflavin Ameliorates Steatotic Liver Ischemia - Reperfusion Injury in MicxGamma AINo ratings yet
- Cell Transplantation-Based Regenerative Medicine in Liver DiseasesDocument18 pagesCell Transplantation-Based Regenerative Medicine in Liver DiseasesalumnogustaNo ratings yet
- Hepatology - 2008 - Yovchev - Identification of Adult Hepatic Progenitor Cells Capable of Repopulating Injured Rat Liver6Document12 pagesHepatology - 2008 - Yovchev - Identification of Adult Hepatic Progenitor Cells Capable of Repopulating Injured Rat Liver6relbelkasyNo ratings yet
- Seminario DCDocument26 pagesSeminario DCHans MenaresNo ratings yet
- Phosphorus Restriction Prevents Parathyroid Gland Growth: High Phosphorus Directly Stimulates PTH Secretion in VitroDocument7 pagesPhosphorus Restriction Prevents Parathyroid Gland Growth: High Phosphorus Directly Stimulates PTH Secretion in VitroDaniel Lee Eisenberg JacobsNo ratings yet
- Resveratrol Attenuates Oxidative Stress and Histological Alterations Induced by Liver Ischemia/reperfusion in RatsDocument6 pagesResveratrol Attenuates Oxidative Stress and Histological Alterations Induced by Liver Ischemia/reperfusion in RatsAbi ZakiNo ratings yet
- Seminario 05 Fibrosis Hepatica Eulenberg 2017Document16 pagesSeminario 05 Fibrosis Hepatica Eulenberg 2017Hello pandiNo ratings yet
- Beyer 1992Document8 pagesBeyer 1992Stress LessNo ratings yet
- 2016.Zheng-Hai - Activating Nuclear Xenobiotic Receptors and Triggering ER Stress and Hepatic Cytochromes P450 Systems in Quails (Coturnix C-Coturnix) During Atrazine ExposureDocument10 pages2016.Zheng-Hai - Activating Nuclear Xenobiotic Receptors and Triggering ER Stress and Hepatic Cytochromes P450 Systems in Quails (Coturnix C-Coturnix) During Atrazine ExposureIasmin ResendeNo ratings yet
- Effect of Quercetin On Paraoxonase 1 Activity - Studies in Cultured Cells, Mice and HumansDocument7 pagesEffect of Quercetin On Paraoxonase 1 Activity - Studies in Cultured Cells, Mice and Humansanisa silviNo ratings yet
- Cutanea Tarda PorphyriaDocument9 pagesCutanea Tarda PorphyriaKanwal RashidNo ratings yet
- Role of Regeneration in Hepatoprotection: Becky TaubDocument26 pagesRole of Regeneration in Hepatoprotection: Becky TaubAgus SuryawanNo ratings yet
- Role of Mesenchymal Stem Cell Therapy in Cisplatin Induced Nephrotoxicity in Adult Albino Rats: Ultrastructural & Biochemical StudyDocument10 pagesRole of Mesenchymal Stem Cell Therapy in Cisplatin Induced Nephrotoxicity in Adult Albino Rats: Ultrastructural & Biochemical StudyNdra SaputraNo ratings yet
- In Uence of Cancer Cachexia On Drug Liver Metabolism and Renal Elimination in RatsDocument8 pagesIn Uence of Cancer Cachexia On Drug Liver Metabolism and Renal Elimination in RatsGregorius HocevarNo ratings yet
- (Doi 10.1089/neu.2014.3497) A. D. Sauerbeck J. L. Laws V. v. Bandaru P. G. Popovich N. J - Spinal Cord Injury Causes Chronic Liver Pathology in RatsDocument11 pages(Doi 10.1089/neu.2014.3497) A. D. Sauerbeck J. L. Laws V. v. Bandaru P. G. Popovich N. J - Spinal Cord Injury Causes Chronic Liver Pathology in RatsDitaIsnainiNo ratings yet
- AscitesDocument1 pageAsciteseditorveterinaryworldNo ratings yet
- PET CT in Patients With Liver Lesions of Different NatureDocument17 pagesPET CT in Patients With Liver Lesions of Different NatureantelioaugustoNo ratings yet
- Vit ADocument8 pagesVit AshandraNo ratings yet
- (03241750 - Acta Medica Bulgarica) The Heparanase Inhibitor (Sulodexide) Decreases Urine Glycosaminoglycan Excretion and Mitigates Functional and Histological Renal Damages in Diabetic RatsDocument6 pages(03241750 - Acta Medica Bulgarica) The Heparanase Inhibitor (Sulodexide) Decreases Urine Glycosaminoglycan Excretion and Mitigates Functional and Histological Renal Damages in Diabetic RatsTeodorNo ratings yet
- Alterations in Peripheral Purinergic and Muscarinic Signaling of Rat Bladder After Long-Term Fructose-Induced Metabolic SyndromeDocument13 pagesAlterations in Peripheral Purinergic and Muscarinic Signaling of Rat Bladder After Long-Term Fructose-Induced Metabolic SyndromeThalia KarampasiNo ratings yet
- PancreotomizadosDocument9 pagesPancreotomizadosCatarina RafaelaNo ratings yet
- Plasma Leptin Levels in Rats With PancreatitisDocument6 pagesPlasma Leptin Levels in Rats With PancreatitisAndykaYayanSetiawanNo ratings yet
- Ecstasy Liver 1996 GutDocument5 pagesEcstasy Liver 1996 GutDaniela Galvez RiveraNo ratings yet
- Regulación Del Metabolismo de Los Eritrocitos Por La InsulinaDocument11 pagesRegulación Del Metabolismo de Los Eritrocitos Por La InsulinaJesus D. Hernandez GuitianNo ratings yet
- Lead Intoxication in A Puppy - A Case Report: E. S, J. P - G, I. K, B. P, B. A, Z. Š - PDocument2 pagesLead Intoxication in A Puppy - A Case Report: E. S, J. P - G, I. K, B. P, B. A, Z. Š - PMauricio RíosNo ratings yet
- Focal Single Intravenous Aminonucleoside: Following Dose PuromycinDocument7 pagesFocal Single Intravenous Aminonucleoside: Following Dose PuromycinFauziana NurhanisahNo ratings yet
- Captopril Alleviates Hypertension-Induced Renal Damage, in Ammation, and NF-KB ActivationDocument9 pagesCaptopril Alleviates Hypertension-Induced Renal Damage, in Ammation, and NF-KB ActivationpipikafiyaNo ratings yet
- Screenshot 2021-10-27 at 18.10.44Document9 pagesScreenshot 2021-10-27 at 18.10.44elok_88No ratings yet
- Seminario 4A Regulacion de La GlicemiaDocument6 pagesSeminario 4A Regulacion de La GlicemiaEdol LopezNo ratings yet
- Wilson 2015Document8 pagesWilson 2015ngvy520201No ratings yet
- Tomita 2014Document9 pagesTomita 2014rahman ijonNo ratings yet
- VI - Joint & Connective Tissue Disorders: RA GoutDocument12 pagesVI - Joint & Connective Tissue Disorders: RA GoutSrinivas SrinuNo ratings yet
- Chronic Metabolic Disorder-Recurrent Acute Arthritis & Inflammation-Na Urate Crystal - Joints HyperurecemiaDocument7 pagesChronic Metabolic Disorder-Recurrent Acute Arthritis & Inflammation-Na Urate Crystal - Joints HyperurecemiaSrinivas SrinuNo ratings yet
- Diabetis 2Document23 pagesDiabetis 2Srinivas SrinuNo ratings yet
- NSAIDs, GOUTDocument18 pagesNSAIDs, GOUTSrinivas SrinuNo ratings yet
- Materia: MedicaDocument650 pagesMateria: MedicaSrinivas SrinuNo ratings yet
- Thyroid 32Document19 pagesThyroid 32Srinivas SrinuNo ratings yet
- M.pharm PCI Syllabus JNTUK MPH R 20 Final BlackDocument227 pagesM.pharm PCI Syllabus JNTUK MPH R 20 Final BlackSrinivas SrinuNo ratings yet
- HomeoDocument34 pagesHomeoSrinivas SrinuNo ratings yet
- This Video Is About How To Download Scribd Documents FreeDocument1 pageThis Video Is About How To Download Scribd Documents FreeSrinivas SrinuNo ratings yet
- Antiulcer METHODOLOGYDocument5 pagesAntiulcer METHODOLOGYSrinivas SrinuNo ratings yet
- How To Download Scribd Documents FreeDocument1 pageHow To Download Scribd Documents FreeSrinivas SrinuNo ratings yet
- Parasitologists United J 2012 5 2 189 198 PDFDocument10 pagesParasitologists United J 2012 5 2 189 198 PDFSrinivas SrinuNo ratings yet
- Antioxidant Activity of Ethanolic Extract of Some Pulp of Fruit of Zizyphus Jujuba Produces in The Algerian West in Vivo PDFDocument5 pagesAntioxidant Activity of Ethanolic Extract of Some Pulp of Fruit of Zizyphus Jujuba Produces in The Algerian West in Vivo PDFSrinivas SrinuNo ratings yet
- Sankaran S Plant Sensations in Pictures Rajan Sankaran Sandra Petri.11046 1contentsDocument3 pagesSankaran S Plant Sensations in Pictures Rajan Sankaran Sandra Petri.11046 1contentsSrinivas SrinuNo ratings yet
- 4-2 Final Hepatoprotective PDFDocument71 pages4-2 Final Hepatoprotective PDFSrinivas SrinuNo ratings yet
- Clinical Surgery - A Practical GuideDocument449 pagesClinical Surgery - A Practical GuideNewvov100% (9)
- MCQ InternalDocument21 pagesMCQ InternalKowsalya SelvarajNo ratings yet
- Goljan Audios Transcribed (Margaret)Document216 pagesGoljan Audios Transcribed (Margaret)sean8phamNo ratings yet
- SopDocument13 pagesSopAnita OctaviaNo ratings yet
- Multidisciplinary Management of Hepatocellular CarcinomaDocument45 pagesMultidisciplinary Management of Hepatocellular CarcinomaSalmanArifNo ratings yet
- Wilson's DiseaseDocument52 pagesWilson's DiseaseAmit KumarNo ratings yet
- 10 Systemic From Liver Necrosis To 2ry Biliary CirrhosisDocument33 pages10 Systemic From Liver Necrosis To 2ry Biliary CirrhosisadelekeyusufNo ratings yet
- All Conditions Data NCTDocument506 pagesAll Conditions Data NCTMuneer AshrafNo ratings yet
- Schistosomiasis Case StudyDocument5 pagesSchistosomiasis Case Studyapi-318749549No ratings yet
- HA Case Study Ms CelesDocument7 pagesHA Case Study Ms CelesJohn FredNo ratings yet
- Week 8 Hepato Biliary 2Document94 pagesWeek 8 Hepato Biliary 2طفوف محمد.No ratings yet
- Iridology - Health Analysis and Treatments From The Iris of The EyeDocument130 pagesIridology - Health Analysis and Treatments From The Iris of The EyeSilviane Silvério100% (1)
- Anatomy and Physiology of The LiverDocument2 pagesAnatomy and Physiology of The LiverAldjen SetiasNo ratings yet
- Body MRIDocument533 pagesBody MRIInes FerreiraNo ratings yet
- 5/ What Is The Relationship Between Structure of The Small Intestine and Its Function?Document3 pages5/ What Is The Relationship Between Structure of The Small Intestine and Its Function?just kimbooNo ratings yet
- MCQ's With KEY Medicine - BDocument8 pagesMCQ's With KEY Medicine - BSiraj Ul IslamNo ratings yet
- BioSignature Review - Are Hormones The Key To Weight LossDocument19 pagesBioSignature Review - Are Hormones The Key To Weight LossVladimir OlefirenkoNo ratings yet
- Lactate PhysiologyDocument5 pagesLactate PhysiologyLee June LyngNo ratings yet
- Anatomy of PeritoneumDocument61 pagesAnatomy of PeritoneumFirdaus SeptiawanNo ratings yet
- JAUNDICE and CHOLECYSTITISDocument109 pagesJAUNDICE and CHOLECYSTITISSarah Racheal AkelloNo ratings yet
- Liver Disease by Rehana MamDocument20 pagesLiver Disease by Rehana Mamraisameh23No ratings yet
- RawPeople - Magnesium StearateDocument2 pagesRawPeople - Magnesium StearategreenbeeNo ratings yet
- Timbrell J.A. - Poison Paradox - How and When Chemicals Are Toxic (2005)Document361 pagesTimbrell J.A. - Poison Paradox - How and When Chemicals Are Toxic (2005)Ronaldo SantosNo ratings yet
- English ProjectDocument126 pagesEnglish ProjectShivam GuptaNo ratings yet
- Liver As Endocrine - Stuart WhiteDocument53 pagesLiver As Endocrine - Stuart WhitechicanahenoNo ratings yet
- Hepatic, Pancreatic, and Rare Gastrointestinal Complications of Immune Checkpoint Inhibitor Therapy - UpToDateDocument29 pagesHepatic, Pancreatic, and Rare Gastrointestinal Complications of Immune Checkpoint Inhibitor Therapy - UpToDatemayteveronica1000No ratings yet
- Urinalysis Results InterpretationDocument21 pagesUrinalysis Results InterpretationSutjipto WijonoNo ratings yet