You might also like
- Water Supply and Urban Drainage PDFDocument107 pagesWater Supply and Urban Drainage PDFManiko Maniko100% (1)
- Super Critical CO2 As Green SolventDocument11 pagesSuper Critical CO2 As Green SolventAnushka ChakrabortyNo ratings yet
- Chapter 3&4Document36 pagesChapter 3&4Màrwå Rägb ZäkyNo ratings yet
- SFE Review: Advantages of Supercritical Fluid Extraction in Food AnalysisDocument5 pagesSFE Review: Advantages of Supercritical Fluid Extraction in Food AnalysisNuradila Athirah LockmanNo ratings yet
- Sisnteticos VisualesDocument10 pagesSisnteticos Visualessamuel jacinto orozcoNo ratings yet
- We Are Intechopen, The World'S Leading Publisher of Open Access Books Built by Scientists, For ScientistsDocument11 pagesWe Are Intechopen, The World'S Leading Publisher of Open Access Books Built by Scientists, For ScientistsAbe SNo ratings yet
- Chapt07 WaterChemistryDocument52 pagesChapt07 WaterChemistryDEEPEST Love ForeverNo ratings yet
- Pursuing Practical Elegance in Chemical Synthesis: Ryoji NoyoriDocument5 pagesPursuing Practical Elegance in Chemical Synthesis: Ryoji NoyoriTran Thanh HaNo ratings yet
- Continuous Salt Precipitation and Separation From Supercritical Water. Part 1Document14 pagesContinuous Salt Precipitation and Separation From Supercritical Water. Part 1Israel Arias GonzálezNo ratings yet
- Science-experiments-guideDocument13 pagesScience-experiments-guidebabiato41No ratings yet
- Svny335 Exp 02 16 27Document13 pagesSvny335 Exp 02 16 27CURTHBERT NDLOVUNo ratings yet
- Supercritical FluidsDocument4 pagesSupercritical FluidsRhea Lyn LamosteNo ratings yet
- The Kinetics of Degradation of Sodium LauroylsarcosinateDocument24 pagesThe Kinetics of Degradation of Sodium LauroylsarcosinateJoe YomiNo ratings yet
- Water Pollution Causes and SolutionsDocument8 pagesWater Pollution Causes and SolutionsJoan254No ratings yet
- Supercritical Water Gasification of Manure A Thermodynamic Equilibrium Modeling ApproachDocument11 pagesSupercritical Water Gasification of Manure A Thermodynamic Equilibrium Modeling ApproachAyush daveNo ratings yet
- Unesco - Eolss Sample Chapters: Food WasteDocument10 pagesUnesco - Eolss Sample Chapters: Food WasteherikNo ratings yet
- CV3015 - Assignment 2 - Yandi Layadi Lay (U1120277F)Document4 pagesCV3015 - Assignment 2 - Yandi Layadi Lay (U1120277F)Yandi LayadiNo ratings yet
- Solubility and DissolutionDocument37 pagesSolubility and DissolutionAdi_Putra_TanayaNo ratings yet
- Topic 4.2 BOD Model and Oxygen Sag CurveDocument17 pagesTopic 4.2 BOD Model and Oxygen Sag CurveMark NalNo ratings yet
- 6.5 Translocation and Transformation Processes14 Heuristics For Hydrolysis Half-Lives and Susceptibility of Contaminants ToDocument24 pages6.5 Translocation and Transformation Processes14 Heuristics For Hydrolysis Half-Lives and Susceptibility of Contaminants TopapsaNo ratings yet
- Chem Project ASWATHDocument22 pagesChem Project ASWATHAswathNo ratings yet
- SOLUBILITY REVIEWDocument37 pagesSOLUBILITY REVIEWBrahma Hakim Yuanda HutabaratNo ratings yet
- Nutrients: Deuterium-Depleted Water Influence On The Isotope H H Regulation in Body and Individual AdaptationDocument19 pagesNutrients: Deuterium-Depleted Water Influence On The Isotope H H Regulation in Body and Individual AdaptationXabier EzkerraNo ratings yet
- Q. Define COD and TOC in Industrial Pollution?Document8 pagesQ. Define COD and TOC in Industrial Pollution?Sudipta kumar muduliNo ratings yet
- UntitledDocument115 pagesUntitledRekha GovindanNo ratings yet
- Hydrogeology1 2 2 3Document19 pagesHydrogeology1 2 2 3lainefrancisco016No ratings yet
- Nutrient Gas KarbonatDocument56 pagesNutrient Gas KarbonatRonawati SilabanNo ratings yet
- CBR Water Genchem Task 1Document19 pagesCBR Water Genchem Task 1Maya JenitaNo ratings yet
- Water Structure and Properties PDFDocument7 pagesWater Structure and Properties PDFfajar_setia100% (2)
- All About Supercritical FluidsDocument5 pagesAll About Supercritical FluidsPaul Christian SantistebanNo ratings yet
- N Muller Relative Permeability TPM 2011Document19 pagesN Muller Relative Permeability TPM 2011Reza OskuiNo ratings yet
- Angew. Chem. Int. Ed. 2005, 44, 2672 - 2692Document21 pagesAngew. Chem. Int. Ed. 2005, 44, 2672 - 2692Sandro Gonzalez AriasNo ratings yet
- Heterogeneous Catalytic Process For Wastewater Treatment: Ting ZhangDocument30 pagesHeterogeneous Catalytic Process For Wastewater Treatment: Ting Zhangjann youngNo ratings yet
- A Review of The Reactivity of OrganicDocument22 pagesA Review of The Reactivity of OrganicMedelky AnouNo ratings yet
- SUPERCRITICAL-FLUIDDocument2 pagesSUPERCRITICAL-FLUIDPaul Christian SantistebanNo ratings yet
- Hydrogen Production From Biomass Wastes by Hydrothermal GasificationDocument15 pagesHydrogen Production From Biomass Wastes by Hydrothermal GasificationHartono PrayitnoNo ratings yet
- Pharmaceutical Solutions and SolubilityDocument14 pagesPharmaceutical Solutions and SolubilityOke OluwafemiNo ratings yet
- Supercritical For SubmitionDocument7 pagesSupercritical For SubmitionPrashant KumarNo ratings yet
- Part I: Introduction To Water Pollution and Control 1. Water Pollution 1.1water PollutantsDocument8 pagesPart I: Introduction To Water Pollution and Control 1. Water Pollution 1.1water PollutantsBenson Mwathi MungaiNo ratings yet
- Physico-Chemical Water Sampling ProtocolDocument5 pagesPhysico-Chemical Water Sampling ProtocolRyan Carlo CondeNo ratings yet
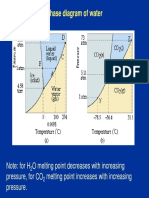
- Phase Diagram of WaterDocument29 pagesPhase Diagram of WaterSaif UllahNo ratings yet
- Aeration and Agitation-ScaleupDocument56 pagesAeration and Agitation-ScaleupVNo ratings yet
- Chapter 4Document50 pagesChapter 4MAHMOUD HAMMADNo ratings yet
- Microbial Decomposition: To Cite This VersionDocument32 pagesMicrobial Decomposition: To Cite This VersionAdarsh KumarNo ratings yet
- Chapter2-WaterDocument32 pagesChapter2-WaterSumia QuashieNo ratings yet
- 22 - Microporous Water With High Gas SolubilitiesDocument24 pages22 - Microporous Water With High Gas SolubilitiesAlex. SonicaNo ratings yet
- Chemical Equilibria in The Surficial EnvironmentDocument34 pagesChemical Equilibria in The Surficial EnvironmentAgnesNo ratings yet
- Water Chemistry Masaka2Document65 pagesWater Chemistry Masaka2Shatho LentsweNo ratings yet
- 2016 - Mosin - Heavy Water As Medium For Living OrganismsDocument27 pages2016 - Mosin - Heavy Water As Medium For Living OrganismsJORGENo ratings yet
- Post Lab and Pre Lab Questions - Cycle-I (2 Files Merged)Document11 pagesPost Lab and Pre Lab Questions - Cycle-I (2 Files Merged)Sayan GharNo ratings yet
- 5SecondaryDocument42 pages5SecondaryFarhan SadikNo ratings yet
- Numerical Simulations of The Thermal Impact of Supercritical CO2 Injection On Chemical Reactivity in A Carbonate Saline Reservoir - Andre - AzaroualDocument45 pagesNumerical Simulations of The Thermal Impact of Supercritical CO2 Injection On Chemical Reactivity in A Carbonate Saline Reservoir - Andre - AzaroualwordindustriesNo ratings yet
- 3) Water Pollution V2 PDFDocument84 pages3) Water Pollution V2 PDFPaanjaisNo ratings yet
- Track Specs 03 - Module 3Document30 pagesTrack Specs 03 - Module 3JatskinesisNo ratings yet
- Phase Diagram of CO2Document10 pagesPhase Diagram of CO2Danveer PrasadNo ratings yet
- Factors Influencing Microbial Activity in Anaerobic BioremediationDocument12 pagesFactors Influencing Microbial Activity in Anaerobic BioremediationGianmarcoNo ratings yet
- Steady Rheological Properties of Rotating Biological Contactor (RBC) SludgeDocument7 pagesSteady Rheological Properties of Rotating Biological Contactor (RBC) SludgeJuan Orlando Loaiza MurilloNo ratings yet
- Anaerobic wastewater treatment processes explained in 38 charactersDocument12 pagesAnaerobic wastewater treatment processes explained in 38 charactersSabhari NatarajanNo ratings yet
- Polymer Processing With Super Critical Fluids by SG KazarianDocument24 pagesPolymer Processing With Super Critical Fluids by SG KazarianLia NashNo ratings yet
- Microbial Ecology Orgainc Aggragates AME2002Document38 pagesMicrobial Ecology Orgainc Aggragates AME2002Mar LunaNo ratings yet
- Báo Cáo Lên Vị Trí - Thung RDDocument2 pagesBáo Cáo Lên Vị Trí - Thung RDNI TomNo ratings yet
- Report FixDocument32 pagesReport FixNI TomNo ratings yet
- Mo HWS13 Dtthung 1513349Document2 pagesMo HWS13 Dtthung 1513349NI TomNo ratings yet
- Down PDFDocument103 pagesDown PDFNI TomNo ratings yet
- Team Project in Reaction With SeparationDocument61 pagesTeam Project in Reaction With SeparationNI TomNo ratings yet
- Numberone PDFDocument15 pagesNumberone PDFNI TomNo ratings yet
- Binary Phase Diagrams: Viet Nam University Ho Chi Minh City HCM University TechnologyDocument11 pagesBinary Phase Diagrams: Viet Nam University Ho Chi Minh City HCM University TechnologyNI TomNo ratings yet
- Alker - 2008 - The Nubian Sandstone Aquifer System (NSAS)Document43 pagesAlker - 2008 - The Nubian Sandstone Aquifer System (NSAS)Mcn calmendNo ratings yet
- How Melting Snow and Dams Can Cause Devastating FloodsDocument3 pagesHow Melting Snow and Dams Can Cause Devastating FloodscalebNo ratings yet
- Technical ManualDocument48 pagesTechnical ManualStewart BenNo ratings yet
- 1204-Barge 2 Tech SpecsDocument13 pages1204-Barge 2 Tech SpecsmardiradNo ratings yet
- SOP 75v4 Specific-Gravity FinalDocument22 pagesSOP 75v4 Specific-Gravity FinalAldwiNo ratings yet
- Packaging N Labelling of Packaged Drinking WaterDocument10 pagesPackaging N Labelling of Packaged Drinking WaterISHFAQ ASHRAFNo ratings yet
- VPL Profile HPDocument2 pagesVPL Profile HPVivek AgarwalNo ratings yet
- Chapter 9 Applied MicrobiologyDocument73 pagesChapter 9 Applied MicrobiologyAsmamaw TesfawNo ratings yet
- Implementing Iot in Underwater Communication Using Li-Fi: S.Shakthivel MuruganDocument7 pagesImplementing Iot in Underwater Communication Using Li-Fi: S.Shakthivel MuruganSsm 001No ratings yet
- ACFrOgCq-liQH4 woWQWjYqVYnW9ifJ5SaweyssbGsl rm5DOvGFnDCyaSS1xLwDBFqGAceZwtAwWbK-Ushw44Zi9TcIwvmdh7qRaGTVKNlhdNEIOBKeQGfkxcDocument11 pagesACFrOgCq-liQH4 woWQWjYqVYnW9ifJ5SaweyssbGsl rm5DOvGFnDCyaSS1xLwDBFqGAceZwtAwWbK-Ushw44Zi9TcIwvmdh7qRaGTVKNlhdNEIOBKeQGfkxcMr JNo ratings yet
- Mishra Sir PDFDocument14 pagesMishra Sir PDFSumit Kumar100% (1)
- How Do Refrigerators WorkDocument33 pagesHow Do Refrigerators Worksushant3240No ratings yet
- Business Continuity Plan Template 24Document20 pagesBusiness Continuity Plan Template 24Paramita AstiNo ratings yet
- Singh, Y.K 2006 - Environmental Science - Cap 1Document11 pagesSingh, Y.K 2006 - Environmental Science - Cap 1Maria Isabel Pérez HernándezNo ratings yet
- Rainwater Harvesting and Artificial Recharge - UnescoDocument22 pagesRainwater Harvesting and Artificial Recharge - UnescoGreen Action Sustainable Technology GroupNo ratings yet
- Particulate Preview Wrap-UpDocument11 pagesParticulate Preview Wrap-UpBunny SmithNo ratings yet
- Comparing BOD Levels Using Permanganometry and Iodometry MethodsDocument4 pagesComparing BOD Levels Using Permanganometry and Iodometry MethodsNia inayahNo ratings yet
- Grade 11 Term 3 PowerpointDocument75 pagesGrade 11 Term 3 PowerpointYONDELA MbandezeloNo ratings yet
- Monsoon Safety PDFDocument6 pagesMonsoon Safety PDFparthaNo ratings yet
- Lacos Sweeping Basin SystemDocument18 pagesLacos Sweeping Basin SystemSanto ENo ratings yet
- International Journal of Biological MacromoleculesDocument10 pagesInternational Journal of Biological MacromoleculesAlejandra Gonzalez RuizNo ratings yet
- Pressure VesselDocument6 pagesPressure VesselEshet ShumetNo ratings yet
- Systematic Study of The Phytoplankton Biodiversity of The Sidi Mhamed Benali Lake of Sidi Bel Abbes 8530Document6 pagesSystematic Study of The Phytoplankton Biodiversity of The Sidi Mhamed Benali Lake of Sidi Bel Abbes 8530Chafik MhamdiaNo ratings yet
- GRADES 1 To 12 Daily Lesson Log: ST ST ST ST STDocument9 pagesGRADES 1 To 12 Daily Lesson Log: ST ST ST ST STYen CradleNo ratings yet
- River Bank 2Document6 pagesRiver Bank 2Ja LuoNo ratings yet
- Dot 16445 DS1 PDFDocument30 pagesDot 16445 DS1 PDFjitenderNo ratings yet
- The Dawn Wall SkillsDocument15 pagesThe Dawn Wall Skillsenas salahNo ratings yet
- Water Quality Monitoring System Using IoT and Machine LearningDocument5 pagesWater Quality Monitoring System Using IoT and Machine LearningBVJNo ratings yet
- Bomba Jockey Vertical SumergibleDocument24 pagesBomba Jockey Vertical SumergibleManuelMartinezNo ratings yet