You might also like
- Plumbing Notes 1 PDFDocument57 pagesPlumbing Notes 1 PDFhoneyvie53% (15)
- TM 10-4930-220-13PDocument133 pagesTM 10-4930-220-13PAdvocateNo ratings yet
- Handbook of Algal Biofuels: Aspects of Cultivation, Conversion, and BiorefineryFrom EverandHandbook of Algal Biofuels: Aspects of Cultivation, Conversion, and BiorefineryMostafa El-SheekhNo ratings yet
- Bioresource TechnologyDocument10 pagesBioresource TechnologyTheresia WinonaNo ratings yet
- Review BiocharDocument10 pagesReview BiocharLeynard NatividadNo ratings yet
- Environmental Pollution: SciencedirectDocument12 pagesEnvironmental Pollution: SciencedirectMonnierNo ratings yet
- Bioresource Technology: A B A C A A ADocument10 pagesBioresource Technology: A B A C A A ARAMA ADITYANo ratings yet
- Anaerobic Co-Digestion of Animal Manures and Lignocellulosic Residues As A Potent Approach For Sustainable Biogas Production PDFDocument15 pagesAnaerobic Co-Digestion of Animal Manures and Lignocellulosic Residues As A Potent Approach For Sustainable Biogas Production PDFLAURA DANIELA CARDONA ACUNANo ratings yet
- Baryla, Bernat, SartowskaDocument7 pagesBaryla, Bernat, SartowskaPaula RotaviskyNo ratings yet
- 1 s2.0 S0147651315002080 MainDocument5 pages1 s2.0 S0147651315002080 MainpurbadreeNo ratings yet
- Accepted Manuscript: Bioresource TechnologyDocument32 pagesAccepted Manuscript: Bioresource TechnologyJulio MoralesNo ratings yet
- Analysis of The Biogas Productivity From Dry Anaerobic Digestion of Organic Fraction of Municipal Solid WasteDocument7 pagesAnalysis of The Biogas Productivity From Dry Anaerobic Digestion of Organic Fraction of Municipal Solid WastePrakash AryalNo ratings yet
- Waste ManagementDocument9 pagesWaste ManagementKevin PaterninaNo ratings yet
- 1 s2.0 S0961953421001562 MainDocument13 pages1 s2.0 S0961953421001562 MainDavid AfricanoNo ratings yet
- Effects of Organic Loading Rate On Biogas Production From MacroalgaeDocument29 pagesEffects of Organic Loading Rate On Biogas Production From MacroalgaeNadjla ChaibNo ratings yet
- Journal of Environmental Management: Research ArticleDocument8 pagesJournal of Environmental Management: Research ArticleThobiasNo ratings yet
- Enhanced Volatile Fatty Acids Production During Anaerobic DigestionDocument7 pagesEnhanced Volatile Fatty Acids Production During Anaerobic DigestionHenry VilchezNo ratings yet
- Materials Today: Proceedings: Vijin Prabhu A., Sivaram A.R., Prabhu N., Sundaramahalingam ADocument6 pagesMaterials Today: Proceedings: Vijin Prabhu A., Sivaram A.R., Prabhu N., Sundaramahalingam ARadhwan AbbasNo ratings yet
- African Journal of Biotechnology Vol. 8 (2)Document6 pagesAfrican Journal of Biotechnology Vol. 8 (2)Perpustakaan UPI FABIO UNSOEDNo ratings yet
- A Review of Biogas Production From Palm Oil Mill Effluents Using Different Configurations of BioreactorsDocument12 pagesA Review of Biogas Production From Palm Oil Mill Effluents Using Different Configurations of BioreactorsAli SmewNo ratings yet
- Agua 2Document6 pagesAgua 2MillerCasTroNo ratings yet
- Pirolisis Por MicroondasDocument7 pagesPirolisis Por MicroondasJulianaGomezNo ratings yet
- 1 PBDocument6 pages1 PBvalentinaNo ratings yet
- Steam Pretreatment of Rice HullsDocument10 pagesSteam Pretreatment of Rice HullsChristian DennisNo ratings yet
- Adding Granular Activated Carbon Into Anaerobic Sludge Digestion To Methane Production and Sludge DecompositionDocument8 pagesAdding Granular Activated Carbon Into Anaerobic Sludge Digestion To Methane Production and Sludge DecompositionJoël KoffiNo ratings yet
- XU - 2019 - Hydrogen and Methane Production by Co-Digesting Liquid Swine Manure andDocument8 pagesXU - 2019 - Hydrogen and Methane Production by Co-Digesting Liquid Swine Manure andThobiasNo ratings yet
- Hua Jie Wang 2018Document7 pagesHua Jie Wang 2018datinovNo ratings yet
- Biogas Two Stage With DiagramDocument10 pagesBiogas Two Stage With DiagramalonsoNo ratings yet
- Biogas Sisa Makanan 5Document11 pagesBiogas Sisa Makanan 5Sherin Novia Dwi PareraNo ratings yet
- GCB Bioenergy - 2014 - Yuan - Mass Balance and Life Cycle Assessment of Biodiesel From Microalgae Incorporated WithDocument15 pagesGCB Bioenergy - 2014 - Yuan - Mass Balance and Life Cycle Assessment of Biodiesel From Microalgae Incorporated WithitbwngNo ratings yet
- Biore Source Technology: Shih-Hsin Ho, Shu-Wen Huang, Chun-Yen Chen, Tomohisa Hasunuma, Akihiko Kondo, Jo-Shu ChangDocument8 pagesBiore Source Technology: Shih-Hsin Ho, Shu-Wen Huang, Chun-Yen Chen, Tomohisa Hasunuma, Akihiko Kondo, Jo-Shu ChangViona WidyaNo ratings yet
- Comparing Activated Carbon of Different Particle Sizes On Enhancing Methane Generation in Upflow Anaerobic Digester - XuDocument7 pagesComparing Activated Carbon of Different Particle Sizes On Enhancing Methane Generation in Upflow Anaerobic Digester - XuAn GonNo ratings yet
- Bioresource Technology: Victoria O. Adesanya, Erasmo Cadena, Stuart A. Scott, Alison G. SmithDocument13 pagesBioresource Technology: Victoria O. Adesanya, Erasmo Cadena, Stuart A. Scott, Alison G. Smithnaturally youNo ratings yet
- Bioresource TechnologyDocument15 pagesBioresource TechnologyAmit Kumar ChaurasiaNo ratings yet
- Mohamed Et Al 2017Document9 pagesMohamed Et Al 2017David Levi QuirozNo ratings yet
- Critical Parametric Influence On Microalgae Cultivation Towards 2016 RenewabDocument8 pagesCritical Parametric Influence On Microalgae Cultivation Towards 2016 RenewabAna Claudia AlmeidaNo ratings yet
- Bioresource Technology 273 (2019) 25-33Document9 pagesBioresource Technology 273 (2019) 25-33UnknownNo ratings yet
- 1 Ana - Recuperación de Proteinas (Hidrolisis Alcalina y Precipitacion Acida)Document9 pages1 Ana - Recuperación de Proteinas (Hidrolisis Alcalina y Precipitacion Acida)Elena Rojo de BenitoNo ratings yet
- Artigo 3Document11 pagesArtigo 3Daniel MottaNo ratings yet
- The Direct Pyrolysis and Catalytic Pyrolysis of Nannochloropsis Sp. Residue For Renewable Bio-OilsDocument8 pagesThe Direct Pyrolysis and Catalytic Pyrolysis of Nannochloropsis Sp. Residue For Renewable Bio-OilsYohn Palomino BellidoNo ratings yet
- Sequencing Batch Membrane Photobioreactor For Real Secondary Effluent Polishing Using Native Microalgae Process Performance and Full-Scale ProjectionDocument8 pagesSequencing Batch Membrane Photobioreactor For Real Secondary Effluent Polishing Using Native Microalgae Process Performance and Full-Scale Projectionroilbilad130No ratings yet
- Characterization and Application of Cellulose Acetate Synthesized FromDocument10 pagesCharacterization and Application of Cellulose Acetate Synthesized FromMuhammad Yusuf SaragihNo ratings yet
- 1 Li2011Document7 pages1 Li2011Wafa BoudayaNo ratings yet
- Tan 2017Document11 pagesTan 2017Aysu UlusalNo ratings yet
- Potential Use EnergyDocument6 pagesPotential Use EnergyAlejandra LopezNo ratings yet
- Zhang 2020Document8 pagesZhang 2020GurusangmeshHiremathNo ratings yet
- Experimental Investigation On Sludge Conditioning and D - 2024 - Journal of EnviDocument11 pagesExperimental Investigation On Sludge Conditioning and D - 2024 - Journal of EnviDana MateiNo ratings yet
- PyroDocument9 pagesPyroFA MonsterNo ratings yet
- Waste Management: Golnaz Arab, Daryl MccartneyDocument11 pagesWaste Management: Golnaz Arab, Daryl Mccartneybelkira69No ratings yet
- Bioresource Technology: Y. Arij, S. Fatihah, A.R. RakmiDocument8 pagesBioresource Technology: Y. Arij, S. Fatihah, A.R. RakmiRyan PasupathyNo ratings yet
- Waste Management: Zhihua Chen, Mian Hu, Baihui Cui, Shiming Liu, Dabin Guo, Bo XiaoDocument6 pagesWaste Management: Zhihua Chen, Mian Hu, Baihui Cui, Shiming Liu, Dabin Guo, Bo XiaoCarlos AlvarezNo ratings yet
- Anaerobic Digested Dairy Manure As A Nutrient Supplement For Cultivation of Oil-Rich Green Microalgae Chlorella Sp.Document6 pagesAnaerobic Digested Dairy Manure As A Nutrient Supplement For Cultivation of Oil-Rich Green Microalgae Chlorella Sp.kayeNo ratings yet
- La Biomasa Microbiana y La Abundancia de Genes de Bacterias Oxidantes de Amoníaco Durante El Compostaje de BiosólidosDocument10 pagesLa Biomasa Microbiana y La Abundancia de Genes de Bacterias Oxidantes de Amoníaco Durante El Compostaje de BiosólidosMartha Ximena Ninco CastañedaNo ratings yet
- Bioresource Technology: Sami Luste, Sari LuostarinenDocument8 pagesBioresource Technology: Sami Luste, Sari LuostarinenJim TsikasNo ratings yet
- Optimization of Biogas Production Through Selection of Appropriate Inoculum-to-Substrate ( (I/S) RatioDocument8 pagesOptimization of Biogas Production Through Selection of Appropriate Inoculum-to-Substrate ( (I/S) RatioSamuel JosephNo ratings yet
- 1 s2.0 S0959652620358212 MainDocument11 pages1 s2.0 S0959652620358212 MainTatiana Izato AssisNo ratings yet
- A Biorefinery Concept Using The Green MacroalgaeDocument9 pagesA Biorefinery Concept Using The Green MacroalgaeNadjla ChaibNo ratings yet
- Chemical Engineering Journal: Rocío Montañés, Montserrat Pérez, Rosario SoleraDocument8 pagesChemical Engineering Journal: Rocío Montañés, Montserrat Pérez, Rosario SoleraMiguel Angel Ramos RuizNo ratings yet
- Biohythane Production From Microalgae-WastewaterDocument23 pagesBiohythane Production From Microalgae-Wastewatergasparperes922No ratings yet
- International Biodeterioration & BiodegradationDocument8 pagesInternational Biodeterioration & BiodegradationArif HidayatNo ratings yet
- Processing Anaerobic Sludge For Extended Storage As Anaerobic Digester InoculumDocument10 pagesProcessing Anaerobic Sludge For Extended Storage As Anaerobic Digester InoculumPizattos SANo ratings yet
- Bioresource Technology: Ke Zhang, Zhijian Pei, Donghai WangDocument13 pagesBioresource Technology: Ke Zhang, Zhijian Pei, Donghai WangAna Sofia Rojas CarpioNo ratings yet
- Bioresource TechnologyDocument8 pagesBioresource TechnologytaniaNo ratings yet
- Bioresource TechnologyDocument8 pagesBioresource TechnologytaniaNo ratings yet
- Bioresource TechnologyDocument8 pagesBioresource TechnologytaniaNo ratings yet
- GalvanGonzalezCohenAlonaizanChenRichSlots Sodium Hypochlorite Oral RinseDocument7 pagesGalvanGonzalezCohenAlonaizanChenRichSlots Sodium Hypochlorite Oral RinsetaniaNo ratings yet
- Oleic Acid TechnologiesDocument11 pagesOleic Acid Technologiessoham kawaleNo ratings yet
- Antinociceptive Activity of Buddleja Globosa (Matico)Document6 pagesAntinociceptive Activity of Buddleja Globosa (Matico)alinumlNo ratings yet
- Who We Are: Case Study-Mahindra ChlorisDocument25 pagesWho We Are: Case Study-Mahindra ChlorisSUNIDHI VERMANo ratings yet
- MasterSpec Consolidated TOCDocument39 pagesMasterSpec Consolidated TOCsheevesNo ratings yet
- 51314-3985-Methanol-Induced Internal Stress CorrosDocument18 pages51314-3985-Methanol-Induced Internal Stress CorrosMahmoud GamalNo ratings yet
- Al Techbull Al195Document2 pagesAl Techbull Al195Ian OttoNo ratings yet
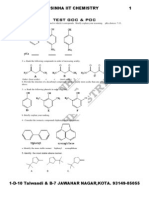
- Test1 Goc & Poc Tough by S.K.sinha See Chemistry Animations atDocument3 pagesTest1 Goc & Poc Tough by S.K.sinha See Chemistry Animations atmyiitchemistry100% (1)
- Hagglunds CaDocument19 pagesHagglunds CaJonathan Giraldo100% (1)
- CRC 2-26Document2 pagesCRC 2-26saranNo ratings yet
- Cambridge International Examinations Cambridge International Advanced Subsidiary and Advanced LevelDocument20 pagesCambridge International Examinations Cambridge International Advanced Subsidiary and Advanced LevelharshanauocNo ratings yet
- Handling of Hygroscopic Products System-TechnikDocument4 pagesHandling of Hygroscopic Products System-TechnikMudassir FarooqNo ratings yet
- TGA Supersoil 5lbDocument1 pageTGA Supersoil 5lbKatherine CampbellNo ratings yet
- Poisoning in ChildrenDocument46 pagesPoisoning in ChildrenpediatricsNo ratings yet
- Freecor LPC English 0Document7 pagesFreecor LPC English 0mgamal1080No ratings yet
- Project Report On Pigment Print BinderDocument4 pagesProject Report On Pigment Print BinderEIRI Board of Consultants and PublishersNo ratings yet
- Application of ESP For Gas Cleaning in Cement Industry - With Reference To IndiaDocument24 pagesApplication of ESP For Gas Cleaning in Cement Industry - With Reference To IndiaSJ ChuaNo ratings yet
- Eisco Industrial Catalogue - FinalDocument276 pagesEisco Industrial Catalogue - FinalSOHAM AlhatNo ratings yet
- Multiple Choice Questions (MCQ) Topic Quiz Biological MembranesDocument22 pagesMultiple Choice Questions (MCQ) Topic Quiz Biological MembranesVeluz Kirt Peter GuiribaNo ratings yet
- A Rapid Accurate Method For The Analysis of An Iron OreDocument5 pagesA Rapid Accurate Method For The Analysis of An Iron Oreilham_metallurgy6744No ratings yet
- UO-II Chapter 4 (Adsorption)Document27 pagesUO-II Chapter 4 (Adsorption)Ghaydah Hamed Rashid Al-AbriNo ratings yet
- Biological Nitrogen FixationDocument24 pagesBiological Nitrogen FixationRam Nivas Ahirwar100% (2)
- Microscope: Trace Evidence PowerpointDocument17 pagesMicroscope: Trace Evidence PowerpointCarlos A. Aponte OteroNo ratings yet
- (En GB) DBE 821 - 1.00Document7 pages(En GB) DBE 821 - 1.00Enio Miguel Cano LimaNo ratings yet
- Exercises: Not One of The Possible Answers ListedDocument12 pagesExercises: Not One of The Possible Answers ListedSarah ChoiNo ratings yet
- Extraccion 4 PDFDocument9 pagesExtraccion 4 PDFsergioenriquerozoperNo ratings yet
- LBR 2014 - ReDocument62 pagesLBR 2014 - ReEugen XhaferajNo ratings yet
- Cbse Class 10 Science SyllabusDocument2 pagesCbse Class 10 Science Syllabusराहुल रविराजNo ratings yet