You might also like
- Ensayos InmunológicosDocument18 pagesEnsayos InmunológicosJorge Luis Olivar100% (1)
- UD2. Técnicas Inmunológicas Basadas en Reacciones Antígeno Anticuerpo Secundarias o IndirectasDocument24 pagesUD2. Técnicas Inmunológicas Basadas en Reacciones Antígeno Anticuerpo Secundarias o IndirectasVerónicaNo ratings yet
- Antígenos y AnticuerposDocument44 pagesAntígenos y AnticuerposAbby Jurz0% (1)
- Actividades Tema 4 - JoseAPintoFDocument10 pagesActividades Tema 4 - JoseAPintoFJOSE ANTONIO PINTORNo ratings yet
- Unidad 3 Reacciones Antigeno-AnticuerpoDocument24 pagesUnidad 3 Reacciones Antigeno-AnticuerpoIsabella Van Buuren100% (1)
- Inmunoglobulinas PDFDocument23 pagesInmunoglobulinas PDFClaudia Patricia Ballen GarzonNo ratings yet
- Métodos de Análisis InmunológicosDocument21 pagesMétodos de Análisis InmunológicosMartin VilledaNo ratings yet
- Inmunología Clase 3: ¿Qué Es Un ANTÍGENO? Molécula/microorganismo/sustancia Que El Organismo DetectaDocument16 pagesInmunología Clase 3: ¿Qué Es Un ANTÍGENO? Molécula/microorganismo/sustancia Que El Organismo DetectaAlfonsina RocciaNo ratings yet
- Inmuno Actividad 3Document8 pagesInmuno Actividad 3ninoska maldonado diazNo ratings yet
- Reacc Antig AnticDocument5 pagesReacc Antig AnticEvelyn RodriguezNo ratings yet
- Guia 2 UcmcDocument7 pagesGuia 2 UcmcSANTIAGO ANDR�S VARGAS MART�NEZNo ratings yet
- Manual de Diagnostico UCMCDocument104 pagesManual de Diagnostico UCMCGeraldine RojasNo ratings yet
- InmunohematologiaDocument27 pagesInmunohematologiaLucila ViqueNo ratings yet
- Metodos IEFDocument48 pagesMetodos IEFmaria del carmenNo ratings yet
- Serologia - Quimica ClinicaDocument17 pagesSerologia - Quimica Clinicaeduardo92% (12)
- INMUNOHISTOQUÍMICADocument39 pagesINMUNOHISTOQUÍMICADarling Escobar UretaNo ratings yet
- Tópicos Básicos Sobre El InmunoensayoDocument7 pagesTópicos Básicos Sobre El InmunoensayoFerney Muñetón AlvarezNo ratings yet
- Tema 5. Obtención y Producción de AnticuerposDocument13 pagesTema 5. Obtención y Producción de AnticuerposSofía Parra ScafiezzoNo ratings yet
- Guia de Discusión 4Document7 pagesGuia de Discusión 4Dove CameronNo ratings yet
- INMUNOLOGIA Tecnicas de LaboratorioDocument32 pagesINMUNOLOGIA Tecnicas de LaboratorioNeelsen EspinolaNo ratings yet
- TP2 y 3Document12 pagesTP2 y 3kewin_walterNo ratings yet
- (Inmunologia) Resumen Teorico 14Document3 pages(Inmunologia) Resumen Teorico 14Samir Torres ChauNo ratings yet
- Inmunidad HumoralDocument7 pagesInmunidad HumoralJavier CeballosNo ratings yet
- Anticuerpos y Complejo Principal de HistocompatibilidadDocument6 pagesAnticuerpos y Complejo Principal de HistocompatibilidadVal OlivaresNo ratings yet
- Tema 6 Introducción A Los Métodos Inmunoquímicos PDFDocument29 pagesTema 6 Introducción A Los Métodos Inmunoquímicos PDFromerillo100% (3)
- Actividad 9. Objetivos BloomDocument6 pagesActividad 9. Objetivos BloomIsaac Marino Cerón AguirreNo ratings yet
- Antigenos e InmunogenosDocument71 pagesAntigenos e InmunogenosMarcovaldo AstolfoNo ratings yet
- Manual Teoricopráctica de InmunologíaDocument32 pagesManual Teoricopráctica de Inmunologíayelenabatista2No ratings yet
- Anticuerpos 2017 PDFDocument53 pagesAnticuerpos 2017 PDFAnchundia Kevin100% (1)
- INMUNOHISTOQUÍMICADocument32 pagesINMUNOHISTOQUÍMICAJulio Mariano Carballo FernandezNo ratings yet
- S2-1 ANTIGENOS y ANTICUERPOS II UD Inmunología y Serodiagnóstico IIDocument64 pagesS2-1 ANTIGENOS y ANTICUERPOS II UD Inmunología y Serodiagnóstico IIyesebellNo ratings yet
- Cuestionario N°3 AnticuerposDocument5 pagesCuestionario N°3 AnticuerposMaria RadcliffeNo ratings yet
- Resumen Ratledge - Cap 25Document12 pagesResumen Ratledge - Cap 25Andres Lopez GuillenNo ratings yet
- Informes Inmunología Semana 9-14Document11 pagesInformes Inmunología Semana 9-14Escoly AltamiranoNo ratings yet
- RX Antigeno-AnticuerpoDocument12 pagesRX Antigeno-AnticuerpoDjvans Samanamud OsorioNo ratings yet
- Reacciones Antigeno Anticuerpo PDFDocument4 pagesReacciones Antigeno Anticuerpo PDFGuadalupe GomezNo ratings yet
- Actividades Unidad 2Document2 pagesActividades Unidad 2CarolinaNo ratings yet
- Antigeno AnticuerpoDocument10 pagesAntigeno AnticuerpoAnnette RamosNo ratings yet
- Ag AcDocument23 pagesAg AcIdamelis Rodríguez GarcíaNo ratings yet
- Grup 6 Imunogloblinas-1Document31 pagesGrup 6 Imunogloblinas-1alisson guayllaNo ratings yet
- Reacciones Ag-AcDocument3 pagesReacciones Ag-AcKaren NCNo ratings yet
- Antigenos y Anticuerpos - Metod - de EstudioDocument30 pagesAntigenos y Anticuerpos - Metod - de EstudioAiser BezaNo ratings yet
- Practica 1Document12 pagesPractica 1Julitaa MazariegosNo ratings yet
- Practica de Reacciones FebrilesDocument8 pagesPractica de Reacciones FebrilesLaura Rosendo CarrilloNo ratings yet
- Antã - Genos y AnticuerposDocument22 pagesAntã - Genos y AnticuerposEDER SOLIS SALDAÑANo ratings yet
- Resumen U2 InmunoDocument11 pagesResumen U2 Inmunolaura chevarria saraviaNo ratings yet
- T.H Antiguenos. AnticuerposDocument5 pagesT.H Antiguenos. AnticuerposAlina ArellanoNo ratings yet
- 6 TECNICAS DE DIAGNÓSTICO LAB. Antígenos - AnticuerposDocument99 pages6 TECNICAS DE DIAGNÓSTICO LAB. Antígenos - AnticuerposNelson CariNo ratings yet
- Inmunologia Tecnicas (Final)Document20 pagesInmunologia Tecnicas (Final)Xiomara AcuñaNo ratings yet
- 06 - Clase Interaccion Ag AcDocument101 pages06 - Clase Interaccion Ag AcdannymalcacamposNo ratings yet
- Cuaderno de Lab. RIA, BLOT Y PCR.Document12 pagesCuaderno de Lab. RIA, BLOT Y PCR.anjiNo ratings yet
- InmunohematologiaDocument6 pagesInmunohematologiaCarla RamelloNo ratings yet
- Antigenos y Anticuerpos Cap 5 InmunoDocument30 pagesAntigenos y Anticuerpos Cap 5 InmunoErika VelásquezNo ratings yet
- Reaccion Ag AcDocument7 pagesReaccion Ag Acmarco samanamud osorioNo ratings yet
- Acceso a Universidad para Mayores de 25 años. Biología 2013-2017.: Solucionario Pruebas 2013-2017From EverandAcceso a Universidad para Mayores de 25 años. Biología 2013-2017.: Solucionario Pruebas 2013-2017No ratings yet
- Transferencia de inmunidad en el binomio madre-cría: Estudio comparativo entre especiesFrom EverandTransferencia de inmunidad en el binomio madre-cría: Estudio comparativo entre especiesNo ratings yet
- Introducción a la Biología: RESÚMENES UNIVERSITARIOSFrom EverandIntroducción a la Biología: RESÚMENES UNIVERSITARIOSRating: 5 out of 5 stars5/5 (1)
- Sistemas Rotatorios en EndodonciaDocument24 pagesSistemas Rotatorios en Endodonciaajcj100% (1)
- Animal PakDocument4 pagesAnimal Pakryuzaki1234No ratings yet
- Programa Fisiología Médica 2017aDocument19 pagesPrograma Fisiología Médica 2017aDiego Meza RyzNo ratings yet
- Monografía de RotavirusDocument9 pagesMonografía de Rotavirusnaldycita0% (2)
- Amador CernudaDocument5 pagesAmador CernudaurumediaNo ratings yet
- Fagocitosis in VivoDocument11 pagesFagocitosis in VivoRodriguez Quiñones Heydi67% (3)
- DebriefingDocument4 pagesDebriefingMara ClarkNo ratings yet
- AREAS FUNDAMENTALES de LA SALUD PÚBLICA La Salud Pública Comprende Cuatro Áreas FundamentalesDocument2 pagesAREAS FUNDAMENTALES de LA SALUD PÚBLICA La Salud Pública Comprende Cuatro Áreas FundamentalesMaría Alejandra Cristina Medina Arias75% (4)
- Guia Artritis - Osteomielitis Las OkDocument33 pagesGuia Artritis - Osteomielitis Las OkYemacadi Casas100% (1)
- SDGDGDF PDFDocument74 pagesSDGDGDF PDFJada DennisNo ratings yet
- Diagnostico Sexualidad 7° y 8°Document3 pagesDiagnostico Sexualidad 7° y 8°Carolina Patricia Hita SierraNo ratings yet
- Diversidad CulturalDocument34 pagesDiversidad CulturalAnonymous 38JYpKgE1No ratings yet
- ReporteEscolaridad DOC4760747 (2140823) PDFDocument5 pagesReporteEscolaridad DOC4760747 (2140823) PDFTiago TassanoNo ratings yet
- Formulario de AfiliacionDocument1 pageFormulario de AfiliacionSEGURITA SST SEGURIDAD Y SALUD EN EL TRABAJONo ratings yet
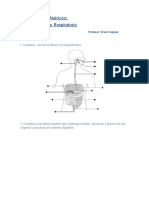
- Nutricón: Sistema Respiratorio: 1. Completa Con Los Nombres CorrespondientesDocument4 pagesNutricón: Sistema Respiratorio: 1. Completa Con Los Nombres CorrespondientesSusana HCNo ratings yet
- Enfermedades Crónicas GhatakDocument299 pagesEnfermedades Crónicas GhatakElizabeth Enriquez100% (9)
- Pruebas VestibularesDocument20 pagesPruebas VestibularesCarolina Cubillos100% (2)
- CorioamnionitisDocument4 pagesCorioamnionitisBae Su JiNo ratings yet
- Acuerdo Ministerial para Conformar Comites de Calidad PDFDocument34 pagesAcuerdo Ministerial para Conformar Comites de Calidad PDFJxzmin Elizxbeth70% (10)
- Anatomía Tercero de Secundaria 06-12-19Document6 pagesAnatomía Tercero de Secundaria 06-12-19Jhon VillaNo ratings yet
- Departamento de Medicamentos Veterinarios: F-DMV-01-12Document5 pagesDepartamento de Medicamentos Veterinarios: F-DMV-01-12rominaNo ratings yet
- Plan Salud Ocupacional PevacDocument34 pagesPlan Salud Ocupacional PevacDïäNäFändïñöNo ratings yet
- La Chica EinsteinDocument335 pagesLa Chica EinsteinWilliam Daniel Amaya Lopez100% (1)
- Pruebas de CompatibilidadDocument26 pagesPruebas de CompatibilidadFernanda VilchesNo ratings yet
- Técnicas PNLDocument14 pagesTécnicas PNLErwing NavasNo ratings yet
- Formato Reporte de Sospecha de Eventos Adversos A MedicamentosDocument4 pagesFormato Reporte de Sospecha de Eventos Adversos A MedicamentosMakiavelo10100% (2)
- Tumores Del Tercer VentriculoDocument36 pagesTumores Del Tercer VentriculoJose Luis Merma PaniuraNo ratings yet
- San Fernando Municipio - Una Ciudad Que Se Renueva. - Deportes 2023Document1 pageSan Fernando Municipio - Una Ciudad Que Se Renueva. - Deportes 2023Ignacio RuedaNo ratings yet
- El Delicuente Sexual SerialDocument17 pagesEl Delicuente Sexual SerialVíctor Eduardo Silva Muñoz100% (1)
- Dermatología PrehispánicaDocument54 pagesDermatología PrehispánicaOctavio MezaNo ratings yet