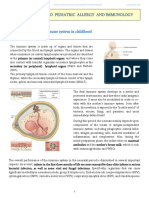
You might also like
- Management of Infections in the Immunocompromised HostFrom EverandManagement of Infections in the Immunocompromised HostBrahm H. SegalNo ratings yet
- Active Immunization Long DOCPDFDocument31 pagesActive Immunization Long DOCPDFEnder IzaguirreNo ratings yet
- Immune System - Development and Acquisition of Immunological Competence - 2020Document8 pagesImmune System - Development and Acquisition of Immunological Competence - 2020Cláudia SilvaNo ratings yet
- The Maturing Immune System: Neonatal Immunity: Table 1Document2 pagesThe Maturing Immune System: Neonatal Immunity: Table 1Vania ParamithaNo ratings yet
- 1 s2.0 S0169409X2030065X MainDocument55 pages1 s2.0 S0169409X2030065X MainLixia WeiNo ratings yet
- Immune Responses in NeonatesDocument26 pagesImmune Responses in NeonatesCony GSNo ratings yet
- Theunis, Louise Immunotherapeutics CapstoneDocument4 pagesTheunis, Louise Immunotherapeutics CapstoneLouise TheunisNo ratings yet
- Basic ImmunologyDocument12 pagesBasic Immunologyjony_phurailatpamNo ratings yet
- TUBE MostCited 1Document6 pagesTUBE MostCited 1jibin2008No ratings yet
- Bases InmunologicasDocument13 pagesBases InmunologicasMadeleine CarmonaNo ratings yet
- Neonatal Immunization Rationale Current State andDocument10 pagesNeonatal Immunization Rationale Current State andNancy CVNo ratings yet
- Vaccine ImmunologyDocument20 pagesVaccine ImmunologyAlejandro MotoliníaNo ratings yet
- Minireview: Neonatal Innate Immunity To Infectious AgentsDocument8 pagesMinireview: Neonatal Innate Immunity To Infectious Agentsapi-3710742No ratings yet
- Rationale For The Immunization Schedule: Whyisitthewayitis?: Education GapsDocument13 pagesRationale For The Immunization Schedule: Whyisitthewayitis?: Education GapsNguyễn Tiến ĐạtNo ratings yet
- Newgen VaccinesDocument7 pagesNewgen Vaccinestranthuytrang213No ratings yet
- VaccinesDocument8 pagesVaccinesArianaNo ratings yet
- Rationale For The Immunization Schedule: Whyisitthewayitis?: Education GapsDocument13 pagesRationale For The Immunization Schedule: Whyisitthewayitis?: Education GapsdelilamhrnNo ratings yet
- Rol de La Inmunidad Innata en La Infección Neonatal. 2013 PDFDocument8 pagesRol de La Inmunidad Innata en La Infección Neonatal. 2013 PDFVictor AyalaNo ratings yet
- Journal of Internal Medicine Volume 253 Issue 4 2003Document9 pagesJournal of Internal Medicine Volume 253 Issue 4 2003vignesh9489No ratings yet
- Pathogenesis of Preterm Birth Bidirectional Inflammation in Mother and FetusDocument17 pagesPathogenesis of Preterm Birth Bidirectional Inflammation in Mother and FetusAlejandro Abarca VargasNo ratings yet
- 1 s2.0 S222116911530366X MainDocument10 pages1 s2.0 S222116911530366X MainNatália FreitasNo ratings yet
- Vaccines 09 00092Document14 pagesVaccines 09 00092Gante FrancesNo ratings yet
- Immunity in Neonates: Bror Morein, Izzeldin Abusugra, Gunilla BlomqvistDocument7 pagesImmunity in Neonates: Bror Morein, Izzeldin Abusugra, Gunilla BlomqvistCatalina Pérez ValderramaNo ratings yet
- Innate Immunity: Fig 1 Table IDocument9 pagesInnate Immunity: Fig 1 Table IforeveraldyNo ratings yet
- Editorial Overview Hildegund CJ Ertl and Antonio LanzavecchiaDocument3 pagesEditorial Overview Hildegund CJ Ertl and Antonio Lanzavecchiahoangan118No ratings yet
- Best Practive Clinical 2021Document17 pagesBest Practive Clinical 2021cdsaludNo ratings yet
- Sistem Imun UptodateDocument21 pagesSistem Imun UptodateAndreea AlexandruNo ratings yet
- Rakib L3 VaccinesDocument23 pagesRakib L3 VaccinesSadia Afrin OshinNo ratings yet
- Working Paper 3 h5n1Document7 pagesWorking Paper 3 h5n1vijayNo ratings yet
- Towards An Epitope-Based Human Vaccine For Influenza: Tamar Ben-Yedidia Ruth ArnonDocument7 pagesTowards An Epitope-Based Human Vaccine For Influenza: Tamar Ben-Yedidia Ruth ArnonPCRMANNo ratings yet
- MF59 Mediates Its B Cell Adjuvanticity by Promoting T Follicular Helper Cells and Thus Germinal Center Responses in Adult and Early LifeDocument11 pagesMF59 Mediates Its B Cell Adjuvanticity by Promoting T Follicular Helper Cells and Thus Germinal Center Responses in Adult and Early LifeJoão Pedro NunesNo ratings yet
- Maternal Antibodies Childhood Infections and AutoimmuneDocument5 pagesMaternal Antibodies Childhood Infections and Autoimmunelempicka23No ratings yet
- Good Cop, Bad Cop? Interrogating The Immune Responses To Primate Lentiviral VaccinesDocument10 pagesGood Cop, Bad Cop? Interrogating The Immune Responses To Primate Lentiviral VaccinesMuhafizNo ratings yet
- Vaccin OlogyDocument79 pagesVaccin Ologyhamzabinamjad987No ratings yet
- Vaccines: Wim Jiskoot, Gideon F. A. Kersten, Enrico Mastrobattista, and Bram SlütterDocument24 pagesVaccines: Wim Jiskoot, Gideon F. A. Kersten, Enrico Mastrobattista, and Bram SlütterSaritaJQM100% (1)
- Vaccination in PregnancyDocument7 pagesVaccination in PregnancyNoraNo ratings yet
- Immunity and Allergy. AsthmaDocument21 pagesImmunity and Allergy. AsthmaSalman SathooNo ratings yet
- Fimmu 04 00407Document12 pagesFimmu 04 00407Alexis LabradaNo ratings yet
- Fimmu 11 00555Document14 pagesFimmu 11 00555Paula VargasNo ratings yet
- Actualidades en InfectologíaDocument47 pagesActualidades en InfectologíaDamián López RangelNo ratings yet
- Antiinfective Properties of Human MilkDocument6 pagesAntiinfective Properties of Human Milkkiko arrojasNo ratings yet
- PIIS0091674900700467Document6 pagesPIIS0091674900700467Ayu DewiNo ratings yet
- Vaccines of The Future Nossal2011Document5 pagesVaccines of The Future Nossal2011Dr Varshil ShahNo ratings yet
- Francis 2015 A Novel Peptide-Based Pan-Influenza A Vaccine RCT of Immunogenicity and SafetyDocument7 pagesFrancis 2015 A Novel Peptide-Based Pan-Influenza A Vaccine RCT of Immunogenicity and SafetyRong LiuNo ratings yet
- DNA VaccinesDocument14 pagesDNA VaccinesTrinity SSRNo ratings yet
- NIH Public Access: Regulatory T Cells and The Immune Pathogenesis of Prenatal InfectionDocument20 pagesNIH Public Access: Regulatory T Cells and The Immune Pathogenesis of Prenatal InfectiongladevaNo ratings yet
- Kenneth, 2009. Sistema Imune SuínoDocument16 pagesKenneth, 2009. Sistema Imune Suínocarol CiveNo ratings yet
- Immunizatuon ChissDocument51 pagesImmunizatuon ChissMariam AntonyNo ratings yet
- 2021 Dawod Et AlDocument10 pages2021 Dawod Et Alpankaj27dataNo ratings yet
- 431 2019 Article 3563Document8 pages431 2019 Article 3563chindiaNo ratings yet
- GIUDICE 2018 - Correlates of Adjuvanticity A Review On Adjuvants in Licensed VaccinesDocument8 pagesGIUDICE 2018 - Correlates of Adjuvanticity A Review On Adjuvants in Licensed VaccinesJoão Pedro NunesNo ratings yet
- Technological Approaches To Streamline Vaccination Schedules, Progressing Towards Single-Dose VaccinesDocument13 pagesTechnological Approaches To Streamline Vaccination Schedules, Progressing Towards Single-Dose VaccinesM OrganicNo ratings yet
- L1 OcrDocument12 pagesL1 OcrMohamed AlsaabNo ratings yet
- Journal Ppat 1000816Document4 pagesJournal Ppat 1000816sanmusikNo ratings yet
- 2015 FonsecaDocument8 pages2015 Fonsecacarlos tNo ratings yet
- New Horizons in Adjuvants For Vaccine Development: Steven G. Reed, Sylvie Bertholet, Rhea N. Coler and Martin FriedeDocument10 pagesNew Horizons in Adjuvants For Vaccine Development: Steven G. Reed, Sylvie Bertholet, Rhea N. Coler and Martin FriedehafsaNo ratings yet
- Ru Ella 2013Document25 pagesRu Ella 2013Ruben NatarîșNo ratings yet
- Microbial Pathogenesis: Reeta Rai, Baijnath Das, Nageshwar Choudhary, Ayantika Talukdar, Donthamsetty Nageswara RaoDocument8 pagesMicrobial Pathogenesis: Reeta Rai, Baijnath Das, Nageshwar Choudhary, Ayantika Talukdar, Donthamsetty Nageswara RaoDiego TulcanNo ratings yet
- VaccinesDocument8 pagesVaccinesRaul MuresanNo ratings yet
- ReviewDocument35 pagesReviewJaganath RaviNo ratings yet
- Strongyloidiasis in HumansDocument11 pagesStrongyloidiasis in HumansLeonardo PalenciaNo ratings yet
- Reduction of Global Cardiovascular Risk With NutritionDocument10 pagesReduction of Global Cardiovascular Risk With NutritionLeonardo PalenciaNo ratings yet
- Estudio EDICDocument10 pagesEstudio EDICLeonardo PalenciaNo ratings yet
- Diagnostic and Prognostic Accuracy of ClinicalDocument10 pagesDiagnostic and Prognostic Accuracy of ClinicalLeonardo PalenciaNo ratings yet
- The Source of MCH Class IDocument6 pagesThe Source of MCH Class ILeonardo PalenciaNo ratings yet
- Acta TropicaDocument5 pagesActa TropicaLeonardo PalenciaNo ratings yet
- 04 SAMSS 005 Check ValvesDocument9 pages04 SAMSS 005 Check ValvesShino UlahannanNo ratings yet
- Kwasaki ZX10R 16Document101 pagesKwasaki ZX10R 16OliverNo ratings yet
- (G. Lakshmi Narasaiah) Finite Element Analysis PDFDocument349 pages(G. Lakshmi Narasaiah) Finite Element Analysis PDFmoljaime1326No ratings yet
- Spectroscopy (L-26 To 29)Document81 pagesSpectroscopy (L-26 To 29)Vashistha GargNo ratings yet
- Illuminati TruthsDocument27 pagesIlluminati TruthsDonnaveo ShermanNo ratings yet
- Active Faults in MalaysiaDocument52 pagesActive Faults in MalaysiaHazim HaNo ratings yet
- Sci NB Mod 2 LSN 3Document4 pagesSci NB Mod 2 LSN 3Ramses octavio Rodriguez ocanasNo ratings yet
- BS746 2014Document22 pagesBS746 2014marco SimonelliNo ratings yet
- Calibration of Force ReductionDocument36 pagesCalibration of Force Reductionvincenzo_12613735No ratings yet
- IPC's 2 Edition of Guidance Manual For Herbs and Herbal Products Monographs ReleasedDocument1 pageIPC's 2 Edition of Guidance Manual For Herbs and Herbal Products Monographs ReleasedRakshaNo ratings yet
- TranscriptDocument1 pageTranscriptapi-310448954No ratings yet
- 09 Passport 7K 15K Performance Guidelines PCR 3 0Document44 pages09 Passport 7K 15K Performance Guidelines PCR 3 0thed719No ratings yet
- Chemical Bonds WorksheetDocument2 pagesChemical Bonds WorksheetJewel Mae MercadoNo ratings yet
- Imbinari Teava Fibra de Sticla PDFDocument1 pageImbinari Teava Fibra de Sticla PDFplex015No ratings yet
- Unnatural Selection BiologyDocument2 pagesUnnatural Selection BiologyAlexa ChaviraNo ratings yet
- Future AncestorsDocument44 pagesFuture AncestorsAlex100% (1)
- ANS: (2.59807m/s2 Horizontal) (1.5m/s2 Vertical) (12.93725 Degree Angle That The Water Surface Makes With The Horizontal)Document5 pagesANS: (2.59807m/s2 Horizontal) (1.5m/s2 Vertical) (12.93725 Degree Angle That The Water Surface Makes With The Horizontal)Lolly UmaliNo ratings yet
- Marketing Management 4th Edition Winer Test BankDocument35 pagesMarketing Management 4th Edition Winer Test Bankapneaocheryzxs3ua100% (24)
- Smoldering Combustion: Guillermo ReinDocument20 pagesSmoldering Combustion: Guillermo ReinAhmed HussainNo ratings yet
- Bhagwati School Strap Report AnalysisDocument60 pagesBhagwati School Strap Report AnalysisReverse Minded100% (1)
- Superposition and Statically Indetermina - GDLCDocument25 pagesSuperposition and Statically Indetermina - GDLCAnonymous frFFmeNo ratings yet
- Yadea User Manual For e Scooter 1546004910Document44 pagesYadea User Manual For e Scooter 1546004910Danthe ThenadNo ratings yet
- Class 12 Maths Project On Prime NumbersDocument13 pagesClass 12 Maths Project On Prime Numbersanon_3835245630% (1)
- Product Stock Exchange Learn BookDocument1 pageProduct Stock Exchange Learn BookSujit MauryaNo ratings yet
- Sch3u Exam Review Ws s2018 PDFDocument4 pagesSch3u Exam Review Ws s2018 PDFwdsfNo ratings yet
- Binet Kamat Test For General Mental AbilitiesDocument54 pagesBinet Kamat Test For General Mental AbilitiesSana Siddiq100% (14)
- Table 1 Minimum Separation DistancesDocument123 pagesTable 1 Minimum Separation DistancesjhonNo ratings yet
- Furuno CA 400Document345 pagesFuruno CA 400Димон100% (3)
- How To Eat WellDocument68 pagesHow To Eat WelleledidiNo ratings yet
- Intel Stratix 10 Avalon - MM Interface For PCI Express Solutions User GuideDocument173 pagesIntel Stratix 10 Avalon - MM Interface For PCI Express Solutions User Guideenoch richardNo ratings yet