You might also like
- Psicobiología UNED - Problemas de Genética RevisadosDocument9 pagesPsicobiología UNED - Problemas de Genética Revisadoscusone100% (3)
- Determinación de Las Consecuencias Que Mantienen La ConductaDocument9 pagesDeterminación de Las Consecuencias Que Mantienen La ConductasergioNo ratings yet
- Ejemplos Reforzamiento Negativo Reforzamiento ParcialDocument7 pagesEjemplos Reforzamiento Negativo Reforzamiento ParcialManuel MamaniNo ratings yet
- Valores y Virtudes MoralesDocument8 pagesValores y Virtudes MoralesLuis AlfonsoNo ratings yet
- Psicología ambientalDocument5 pagesPsicología ambientalAlberto CabelloNo ratings yet
- Actividad 1Document2 pagesActividad 1Scarlet BurgosNo ratings yet
- 1) Ejercicio de Indice de Marco TeoricoDocument2 pages1) Ejercicio de Indice de Marco TeoricoAlex Hernandez SantiagoNo ratings yet
- Actividad 2. Etica ProfesionalDocument2 pagesActividad 2. Etica ProfesionalMaria Teresa CUEVA QUINTERONo ratings yet
- Actividades Didactica PDFDocument7 pagesActividades Didactica PDFEder Julinho Farfan ImanNo ratings yet
- Guia Elaboracion TesisDocument92 pagesGuia Elaboracion TesisGabriel Esteban Beltran ZTNo ratings yet
- Criterios de La Conducta Humana PDFDocument8 pagesCriterios de La Conducta Humana PDFSebastian SandovalNo ratings yet
- ParatextoDocument9 pagesParatextoBeter TresNo ratings yet
- Diseños Cuantitativos PDFDocument38 pagesDiseños Cuantitativos PDFAnonymous 4lpqd6uGLmNo ratings yet
- Proyecto ..Medio AmbienteDocument24 pagesProyecto ..Medio AmbienteNatalyBulejeNo ratings yet
- Importancia de La EpistemologiaDocument12 pagesImportancia de La EpistemologiaBrayan GomezNo ratings yet
- Investigación Documental 1Document14 pagesInvestigación Documental 1sssssssssssssssdddddddNo ratings yet
- Cronograma de Actividades Medio AmbienteDocument5 pagesCronograma de Actividades Medio Ambientearcanghelo3892No ratings yet
- Qué Es El PluralismoDocument10 pagesQué Es El PluralismoJoselyn Lazo MartinezNo ratings yet
- 1.3 Actividad. Paradigmas de La Investigación EducativaDocument4 pages1.3 Actividad. Paradigmas de La Investigación EducativaIleana Berenice PlascenciaNo ratings yet
- Cuaderno de Casos PrácticosDocument15 pagesCuaderno de Casos PrácticosJocelyn DámasoNo ratings yet
- La libertad como destino y acciónDocument4 pagesLa libertad como destino y acciónAlejandra María José100% (1)
- 2-Tipo y Nivel de Investigación-ChenetDocument45 pages2-Tipo y Nivel de Investigación-ChenetAlex QuispealayaNo ratings yet
- Laboratorio DocenteDocument12 pagesLaboratorio DocenteBenigno Gonzalez MaldonadoNo ratings yet
- Habilidades LogicasDocument21 pagesHabilidades LogicasYubiz Tuesta HuablochoNo ratings yet
- Actividad 2Document1 pageActividad 2RUBENNo ratings yet
- Sucesiones y Series Numéricas PDFDocument4 pagesSucesiones y Series Numéricas PDFEl Momero Peruano.No ratings yet
- Violencia Simbólica y Sistema Escolar en Bourdieu PDFDocument11 pagesViolencia Simbólica y Sistema Escolar en Bourdieu PDFMarisaNo ratings yet
- Habilidades Sociales Tema 7Document16 pagesHabilidades Sociales Tema 7wilog25No ratings yet
- Que Es Un Proceso de PensamientoDocument4 pagesQue Es Un Proceso de PensamientoJohana100% (1)
- Actividad 1 ANALISIS DE ARTICULO DE INVESTIGACIONDocument5 pagesActividad 1 ANALISIS DE ARTICULO DE INVESTIGACIONrociopedrazaNo ratings yet
- Taller de Evaluación EducativaDocument4 pagesTaller de Evaluación EducativaettiareNo ratings yet
- Coll-Psicología y Curriculum PDFDocument4 pagesColl-Psicología y Curriculum PDFrane1950513No ratings yet
- Ejercicio Disenos CualitativosDocument5 pagesEjercicio Disenos CualitativosFlor Delgado SanchezNo ratings yet
- Las Teorías de Henri Wallon 2Document5 pagesLas Teorías de Henri Wallon 2Janet Rodríguez GuzmánNo ratings yet
- Educacion PermanenteDocument14 pagesEducacion PermanenteVeronica Jaldin MitaNo ratings yet
- Ejercicios de Psicologia Páginas 10 13Document4 pagesEjercicios de Psicologia Páginas 10 13Valentina Garduño GuerreroNo ratings yet
- Segunda Secuencia Didáctica de EcologíaDocument7 pagesSegunda Secuencia Didáctica de EcologíaGustavo Adolfo García CollmanNo ratings yet
- Pensamiento ModernoDocument12 pagesPensamiento ModernoPsi Claudio Cesar Orias HuayllaNo ratings yet
- Ejemplo SDocument3 pagesEjemplo SraquelNo ratings yet
- Caracteristicas Del Pensamiento FilosoficoDocument11 pagesCaracteristicas Del Pensamiento FilosoficoManuel Medina0% (1)
- Diseños Longitudinales OkDocument10 pagesDiseños Longitudinales OkBetty GomezNo ratings yet
- Secuencia didáctica aguaDocument5 pagesSecuencia didáctica aguaJuan Roman Vazquez RomoNo ratings yet
- Cronograma Actividades ECOLOGÍA Y MEDIO AMBIENTEDocument6 pagesCronograma Actividades ECOLOGÍA Y MEDIO AMBIENTEFausto George Prado DNo ratings yet
- Actividad 2 Paradigmas de La InvestigacionDocument6 pagesActividad 2 Paradigmas de La InvestigacionPOPERSNo ratings yet
- Pensamiento crítico en el aulaDocument26 pagesPensamiento crítico en el aulaDireccion Regional50% (2)
- Proyecto Del Medio AmbienteDocument14 pagesProyecto Del Medio AmbienteLeiner Rios VictorioNo ratings yet
- El Método de Las Ciencias SocialesDocument13 pagesEl Método de Las Ciencias SocialesBelle Medina100% (1)
- Curriculum CognitivoDocument11 pagesCurriculum CognitivoFabiola RamirezNo ratings yet
- Ética y EcologiaDocument11 pagesÉtica y EcologialuisangeloNo ratings yet
- Aplicaciones Biotecnologicas de Los MicroorganismosDocument11 pagesAplicaciones Biotecnologicas de Los MicroorganismosBrayan ChipanaNo ratings yet
- Metodos GravimetricosDocument23 pagesMetodos GravimetricosAli ErnestoNo ratings yet
- Actividad Evaluativa 1 - Diseños de InvestigacionDocument7 pagesActividad Evaluativa 1 - Diseños de InvestigacionDavid VásquezNo ratings yet
- Ensayo. El Método CientíficoDocument3 pagesEnsayo. El Método CientíficoSåntos FernåndoNo ratings yet
- La ReseñaDocument7 pagesLa ReseñaGlëy AlmonteNo ratings yet
- El Paradigma HumanistaDocument8 pagesEl Paradigma HumanistaAlberto Jardón100% (1)
- Secuencia Didáctica Matemàticas 1º GradoDocument6 pagesSecuencia Didáctica Matemàticas 1º GradoAny Michelle Martínez RíosNo ratings yet
- G11 06miguel Moreno MunozDocument22 pagesG11 06miguel Moreno MunozJosue RomeroNo ratings yet
- Actividad #6 Fundamentos Económicos y Sociales para El Desarrollo de La Genética y Su Influencia en El Ser Humano.Document13 pagesActividad #6 Fundamentos Económicos y Sociales para El Desarrollo de La Genética y Su Influencia en El Ser Humano.Luis Villarreal AvalosNo ratings yet
- Biologizacion y Racismo MenendezDocument74 pagesBiologizacion y Racismo MenendezVioleta MontellanoNo ratings yet
- Comportamiento y ActitudesDocument7 pagesComportamiento y ActitudesYamilka A.V.No ratings yet
- Comentarios y Consultas La Psicología Al Servicio de La Humanidad 1979Document6 pagesComentarios y Consultas La Psicología Al Servicio de La Humanidad 1979ramrojo100200No ratings yet
- Galletas para GenaroDocument2 pagesGalletas para GenaroAna M ViloriaNo ratings yet
- La Comida Que Te MoldeaDocument6 pagesLa Comida Que Te MoldeaAna M ViloriaNo ratings yet
- Emociones EspañolDocument3 pagesEmociones EspañolTeacher DanielaNo ratings yet
- Historia de la elaboración de cerveza y vinoDocument7 pagesHistoria de la elaboración de cerveza y vinoAna M ViloriaNo ratings yet
- El Impacto de La Inteligencia Emocional Sobre La Regulación de La EpigenéticaDocument2 pagesEl Impacto de La Inteligencia Emocional Sobre La Regulación de La EpigenéticaAna M ViloriaNo ratings yet
- Aprenderás Los Secretos Del Lenguaje No Hablado Que He Acumulado Durante Más de 15 AñosDocument2 pagesAprenderás Los Secretos Del Lenguaje No Hablado Que He Acumulado Durante Más de 15 AñosAna M ViloriaNo ratings yet
- Romulo GallegosDocument2 pagesRomulo GallegosAna M ViloriaNo ratings yet
- Dieta PaleoDocument9 pagesDieta PaleoAna M ViloriaNo ratings yet
- KaliDocument2 pagesKaliAna M ViloriaNo ratings yet
- Fritz Perls y Claudio Naranjo documentosDocument1 pageFritz Perls y Claudio Naranjo documentosAna M ViloriaNo ratings yet
- Definiendo La BiodescodificaciònDocument4 pagesDefiniendo La BiodescodificaciònAna M ViloriaNo ratings yet
- El Impacto de La Inteligencia Emocional Sobre La Regulación de La EpigenéticaDocument2 pagesEl Impacto de La Inteligencia Emocional Sobre La Regulación de La EpigenéticaAna M ViloriaNo ratings yet
- Tarot OdtDocument2 pagesTarot OdtAna M ViloriaNo ratings yet
- Dibujando El Árbol. Transgeneracional.Document11 pagesDibujando El Árbol. Transgeneracional.Ana M ViloriaNo ratings yet
- Cómo Puede La Dieta Cambiarte El EpigenomaDocument6 pagesCómo Puede La Dieta Cambiarte El EpigenomaAna M ViloriaNo ratings yet
- Cómo Puede La Dieta Cambiarte El EpigenomaDocument6 pagesCómo Puede La Dieta Cambiarte El EpigenomaAna M ViloriaNo ratings yet
- Epigenética y Gestación Por SustituciónDocument18 pagesEpigenética y Gestación Por SustituciónAna M ViloriaNo ratings yet
- Restricción de Crecimiento Fetal, Epigenética y TransmisiónDocument10 pagesRestricción de Crecimiento Fetal, Epigenética y TransmisiónAna M ViloriaNo ratings yet
- Transferencia Epigenética Transgeneracional de La Experiencia TraumáticaDocument6 pagesTransferencia Epigenética Transgeneracional de La Experiencia TraumáticaAna M ViloriaNo ratings yet
- Epigenética en Longevidad y EnvejecimientoDocument6 pagesEpigenética en Longevidad y EnvejecimientoAna M ViloriaNo ratings yet
- La Comida Que Te MoldeaDocument6 pagesLa Comida Que Te MoldeaAna M ViloriaNo ratings yet
- El ARN de Los Espermatozoides Puede Influir en La Herencia de Síntomas Relacionados Con Experiencias TempranasDocument5 pagesEl ARN de Los Espermatozoides Puede Influir en La Herencia de Síntomas Relacionados Con Experiencias TempranasAna M ViloriaNo ratings yet
- Epigenética en Longevidad y EnvejecimientoDocument6 pagesEpigenética en Longevidad y EnvejecimientoAna M ViloriaNo ratings yet
- Modificar El Epigenoma Como Herramienta Frente A La EnfermedadDocument4 pagesModificar El Epigenoma Como Herramienta Frente A La EnfermedadAna M ViloriaNo ratings yet
- Modificar El Epigenoma Como Herramienta Frente A La EnfermedadDocument4 pagesModificar El Epigenoma Como Herramienta Frente A La EnfermedadAna M ViloriaNo ratings yet
- Epigenetica Impronta GenómicaDocument12 pagesEpigenetica Impronta GenómicaAna M ViloriaNo ratings yet
- Epigenetica y Trastornos PsiquiatricosDocument8 pagesEpigenetica y Trastornos PsiquiatricosAna M ViloriaNo ratings yet
- El Estrés Produce Cambios en Los GenesDocument5 pagesEl Estrés Produce Cambios en Los GenesAna M ViloriaNo ratings yet
- Transgeneracional y Epigenética ConductualDocument5 pagesTransgeneracional y Epigenética ConductualAna M ViloriaNo ratings yet
- Las Secuencias y El ParentescoDocument5 pagesLas Secuencias y El ParentescoDaniela MatayoshiNo ratings yet
- Infografía de Los Tipos de Reproducción en Los Seres VivosDocument14 pagesInfografía de Los Tipos de Reproducción en Los Seres VivosDerek ToddNo ratings yet
- El Espacio de Los Escorpiones 1Document53 pagesEl Espacio de Los Escorpiones 1Andrea Ruiz0% (1)
- Pelajeratonesbolsillo Educador Act 0Document8 pagesPelajeratonesbolsillo Educador Act 0Mariela Escobar TorrezNo ratings yet
- CUESTIONARIODocument13 pagesCUESTIONARIOEli MorochoNo ratings yet
- Informe de Acido NucleotidosDocument2 pagesInforme de Acido NucleotidosJorge AlvarezNo ratings yet
- Replicación del ADN y sus etapasDocument93 pagesReplicación del ADN y sus etapasSanti VedoyaNo ratings yet
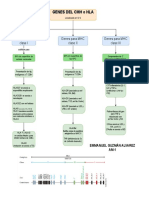
- Genes de HLA o MHCDocument1 pageGenes de HLA o MHCEmmanuel GNo ratings yet
- Guia de Biologia - Preguntas de EvaluaciónDocument99 pagesGuia de Biologia - Preguntas de Evaluaciónvalebu10100% (2)
- Práctica 3. Observación de Células en MeiosisDocument3 pagesPráctica 3. Observación de Células en MeiosissaruausNo ratings yet
- AlphaFold, Inteligencia Artificial para Descubrimiento CientíficoDocument8 pagesAlphaFold, Inteligencia Artificial para Descubrimiento CientíficoantonioavezonNo ratings yet
- Resumen UP2Document11 pagesResumen UP2SofiaNo ratings yet
- La Epigenética Como Consecuencia de La Influencia Del Entorno Medioambiental y Materno en El Desarrollo EmbrionarioDocument10 pagesLa Epigenética Como Consecuencia de La Influencia Del Entorno Medioambiental y Materno en El Desarrollo Embrionariocamila gonzalezNo ratings yet
- 1 Al 36Document35 pages1 Al 36Karen MéndezNo ratings yet
- BM05 22 05 12Document20 pagesBM05 22 05 12feeeleee7No ratings yet
- P2 - Sesión 17Document8 pagesP2 - Sesión 17Ale CortezNo ratings yet
- Avances en La Genetica de La Fisiopatia de La MancDocument2 pagesAvances en La Genetica de La Fisiopatia de La Mancdaniel carrascoNo ratings yet
- TraducciónDocument8 pagesTraducciónMaria FernandaNo ratings yet
- Segundo Turno, Segundo Parcial Teórico-PracticoDocument6 pagesSegundo Turno, Segundo Parcial Teórico-PracticoAna CorreaNo ratings yet
- Practica N 10 - GRUPO 1Document12 pagesPractica N 10 - GRUPO 1erikapasacheNo ratings yet
- 1.teoria Del Envejecimiento-2021.Document34 pages1.teoria Del Envejecimiento-2021.Sarai Hoyos VillavicencioNo ratings yet
- LMAcon Anormalidades GeneticasDocument20 pagesLMAcon Anormalidades GeneticasRudy JumperNo ratings yet
- Vigor HibridoDocument18 pagesVigor HibridoCampos JuanNo ratings yet
- BIOLOGIADocument24 pagesBIOLOGIANohelia CañizalezNo ratings yet
- Nociones de Genética MatematicaDocument71 pagesNociones de Genética MatematicaAugusto Ǝxcelmes Cutimbo100% (1)
- GenéticaDocument3 pagesGenéticaJulián Oliva CáceresNo ratings yet
- 12 Biolog°aDocument8 pages12 Biolog°aEliezer AbregoNo ratings yet
- LA ERA DEL GENOMA (Jesús Purroy) PDFDocument256 pagesLA ERA DEL GENOMA (Jesús Purroy) PDFrodari6147100% (4)