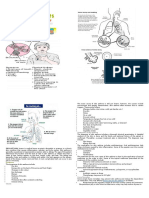
You might also like
- Fichier1038 Phylogeny of Marsilea.Document10 pagesFichier1038 Phylogeny of Marsilea.sellaginellaNo ratings yet
- Murray& ProwellMPE Molecular Phylogenetics and EvolutionaryDocument14 pagesMurray& ProwellMPE Molecular Phylogenetics and EvolutionarysellaginellaNo ratings yet
- 506 Phylogenetic Relationships in Selaginellaceae RBCL +Document12 pages506 Phylogenetic Relationships in Selaginellaceae RBCL +sellaginellaNo ratings yet
- Korall Et Al2 Phylogeny of Selaginellaceae RBCLDocument10 pagesKorall Et Al2 Phylogeny of Selaginellaceae RBCLsellaginellaNo ratings yet
- K - Remarchuk (1) Demography and Genetic Diversity in ADocument136 pagesK - Remarchuk (1) Demography and Genetic Diversity in AsellaginellaNo ratings yet
- Korall Phylogenetic History of Selaginellaceae 26S rDNA + RBCLDocument13 pagesKorall Phylogenetic History of Selaginellaceae 26S rDNA + RBCLsellaginellaNo ratings yet
- 1206 Cardamine Flexuosa (Brassicaceae)Document16 pages1206 Cardamine Flexuosa (Brassicaceae)sellaginellaNo ratings yet
- KJ00000132641 Pteridophytes of Northern Sum - AtraDocument20 pagesKJ00000132641 Pteridophytes of Northern Sum - AtrasellaginellaNo ratings yet
- Fern CpDNA One Particular Taxon Selaginella Was ProblematicDocument14 pagesFern CpDNA One Particular Taxon Selaginella Was ProblematicsellaginellaNo ratings yet
- 271 Chromosome Numbers S.lepidophylaDocument6 pages271 Chromosome Numbers S.lepidophylasellaginellaNo ratings yet
- 506 Phylogenetic Relationships in Selaginellaceae RBCL +Document12 pages506 Phylogenetic Relationships in Selaginellaceae RBCL +sellaginellaNo ratings yet
- S61 Sex Determining MechanismsDocument11 pagesS61 Sex Determining MechanismssellaginellaNo ratings yet
- Phylogenetics in Comparative GeneticsDocument11 pagesPhylogenetics in Comparative GeneticssellaginellaNo ratings yet
- Goodson Et Al. 2006 Biogeographic and Taxonomic ImplicationsDocument12 pagesGoodson Et Al. 2006 Biogeographic and Taxonomic ImplicationssellaginellaNo ratings yet
- 179 Neillieae (Rosaceae)Document14 pages179 Neillieae (Rosaceae)sellaginellaNo ratings yet
- 1230 Lilium Longiflorum+Document10 pages1230 Lilium Longiflorum+sellaginellaNo ratings yet
- Anzfh2dettmann&clifford Biogeography of Araucariaceae+Document9 pagesAnzfh2dettmann&clifford Biogeography of Araucariaceae+sellaginellaNo ratings yet
- 247 Caltha (Ranunculaceae) +Document7 pages247 Caltha (Ranunculaceae) +sellaginellaNo ratings yet
- 601 Fuchsia (Onagraceae) +Document14 pages601 Fuchsia (Onagraceae) +sellaginellaNo ratings yet
- 653 Viburnum Phylogeny+Document14 pages653 Viburnum Phylogeny+sellaginellaNo ratings yet
- Willis and Birks 2006 What Is NaturalDocument5 pagesWillis and Birks 2006 What Is Naturalsellaginella100% (2)
- Triassic LycopsidsDocument35 pagesTriassic LycopsidssellaginellaNo ratings yet
- 217 Platycerium (Polypodiaceae) +Document9 pages217 Platycerium (Polypodiaceae) +sellaginella100% (1)
- 1120 Paeonia (Paeoniaceae)Document17 pages1120 Paeonia (Paeoniaceae)sellaginellaNo ratings yet
- Stepp04 Weeds As Sources of PharmaceuticalsDocument4 pagesStepp04 Weeds As Sources of PharmaceuticalssellaginellaNo ratings yet
- Ogunkunle and Ladejobi Ethnobotanical and Phytochemical StudiesDocument4 pagesOgunkunle and Ladejobi Ethnobotanical and Phytochemical StudiessellaginellaNo ratings yet
- JET - 1999!65!113-124 Interviews Were ConductedDocument12 pagesJET - 1999!65!113-124 Interviews Were ConductedsellaginellaNo ratings yet
- 1387 Keoun BackDocument10 pages1387 Keoun BacksellaginellaNo ratings yet
- Traditional Zulu MedicinalDocument8 pagesTraditional Zulu MedicinalsellaginellaNo ratings yet
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (120)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- Food Allergy Prevention and Treatment - Cow's Milk AllergyDocument19 pagesFood Allergy Prevention and Treatment - Cow's Milk AllergyPatrick nyawiraNo ratings yet
- S 000 LBLDocument14 pagesS 000 LBLfaisal abbasNo ratings yet
- Simman 2024Document54 pagesSimman 2024Doshi SahilNo ratings yet
- Get Trained: A Program For School Nurses To Train School Staff To Administer Epinephrine Using An Auto-Injector © 2015Document34 pagesGet Trained: A Program For School Nurses To Train School Staff To Administer Epinephrine Using An Auto-Injector © 2015Timothy Andre GolsonNo ratings yet
- Adverse Drug Reactions: Rahil Khan H.O.D Clinical Research ICRI MumbaiDocument46 pagesAdverse Drug Reactions: Rahil Khan H.O.D Clinical Research ICRI Mumbairahil_khan797No ratings yet
- Skin AsthmaDocument7 pagesSkin AsthmaJames Domini Lopez LabianoNo ratings yet
- NUR 111 Session 22 SAS SemillaDocument2 pagesNUR 111 Session 22 SAS SemillaCyrille Jhane A GalvezNo ratings yet
- Biology Investigatory ProjectDocument21 pagesBiology Investigatory ProjectCrazy about Jungles87% (30)
- Student Medical FormDocument2 pagesStudent Medical Formapi-351900524No ratings yet
- Anaphylaxis Treatment ProcedureDocument4 pagesAnaphylaxis Treatment ProcedureIvy Jorene Roman RodriguezNo ratings yet
- Definition, Classification, Etiology, and Pathophysiology of Shock in Adults - UpToDateDocument17 pagesDefinition, Classification, Etiology, and Pathophysiology of Shock in Adults - UpToDateSofia Montes Tello100% (14)
- Hypersensitivity Nursing Students (BSC Nursing)Document25 pagesHypersensitivity Nursing Students (BSC Nursing)Ikramul Hussain KhanNo ratings yet
- Wilderness and Remote First Aid: Emergency Reference GuideDocument121 pagesWilderness and Remote First Aid: Emergency Reference GuideAdrian Lindemann100% (1)
- EmergencyDocument86 pagesEmergencyyazzNo ratings yet
- E-Carts Drugs and Their UsesDocument6 pagesE-Carts Drugs and Their UsesMarienelle De La CruzNo ratings yet
- Zosyn Drug CardDocument1 pageZosyn Drug CardSheri490No ratings yet
- Setting The Standard: Managing Food Allergies in SchoolDocument50 pagesSetting The Standard: Managing Food Allergies in SchooldadowildcatsNo ratings yet
- Anaphylaxis EpiPen General Action Plan 2017 WEBDocument1 pageAnaphylaxis EpiPen General Action Plan 2017 WEBDelfi PandieNo ratings yet
- Slemani Pediatric Teaching Hospital GuidelinesDocument93 pagesSlemani Pediatric Teaching Hospital GuidelinesHeersh RaofNo ratings yet
- PICU Protocol 1 Ain Shams UniversityDocument52 pagesPICU Protocol 1 Ain Shams UniversityAhmad ElgazzarNo ratings yet
- Drug Study - HNBBDocument4 pagesDrug Study - HNBBCamilogs100% (3)
- Amicus BriefDocument27 pagesAmicus BriefThe FederalistNo ratings yet
- Cow's Milk Protein AllergyDocument7 pagesCow's Milk Protein Allergy6130015013 MOCHAMMAD SULUNG NOVIYANTO SKMNo ratings yet
- Bepen VKDocument5 pagesBepen VKelcapitano vegetaNo ratings yet
- Anaphylactic Death FinalDocument73 pagesAnaphylactic Death Finalkhaled eissaNo ratings yet
- Homework5 Hazard Research Group#1Document39 pagesHomework5 Hazard Research Group#1Vannya Nayely Hernández SaizaNo ratings yet
- Classification of Drug Hypersensitivity Into Allergic, P-I, and Pseudo-Allergic FormsDocument14 pagesClassification of Drug Hypersensitivity Into Allergic, P-I, and Pseudo-Allergic FormsFanny PritaningrumNo ratings yet
- HypersensitivityDocument15 pagesHypersensitivityTejpalNo ratings yet
- By Mayo Clinic Staff: Milk Allergy or Milk Intolerance?Document5 pagesBy Mayo Clinic Staff: Milk Allergy or Milk Intolerance?riadwiNo ratings yet
- Anaphylaxis: Initial Management in Non-Hospital Settings: Edition", Pages 14 To 18 (Anaphylaxis Section Only)Document4 pagesAnaphylaxis: Initial Management in Non-Hospital Settings: Edition", Pages 14 To 18 (Anaphylaxis Section Only)Fera MulidarNo ratings yet