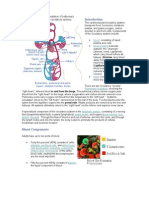
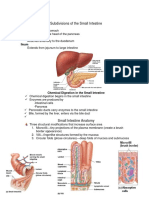
You might also like
- Using Autoethnography in The Scholarship of Teaching and Learning DuarteDocument11 pagesUsing Autoethnography in The Scholarship of Teaching and Learning DuarteRuth Danielle GasconNo ratings yet
- Burns, Metabolism, and Nutritional RequirementsDocument9 pagesBurns, Metabolism, and Nutritional RequirementsRuth Danielle GasconNo ratings yet
- Characteristics of Life QuizDocument2 pagesCharacteristics of Life QuizRuth Danielle GasconNo ratings yet
- Thrombocytopenic Crisis Following BurnsDocument5 pagesThrombocytopenic Crisis Following BurnsRuth Danielle GasconNo ratings yet
- MGNT of The High Risk Pediatric Burn PatientDocument5 pagesMGNT of The High Risk Pediatric Burn PatientRuth Danielle GasconNo ratings yet
- Nutritional Therapy in Major BurnsDocument6 pagesNutritional Therapy in Major BurnsRuth Danielle GasconNo ratings yet
- Pediatric Burn CareDocument4 pagesPediatric Burn CareRuth Danielle GasconNo ratings yet
- 11-Arcs and Central AnglesDocument4 pages11-Arcs and Central AnglesRuth Danielle GasconNo ratings yet
- Chem 40 Assignment 2 For Printing Apr 19Document1 pageChem 40 Assignment 2 For Printing Apr 19Ruth Danielle GasconNo ratings yet
- Course Syllabus and Schedule: College of Science University of The Philippines Diliman, Quezon City 1101Document4 pagesCourse Syllabus and Schedule: College of Science University of The Philippines Diliman, Quezon City 1101Ruth Danielle Gascon0% (1)
- Activity 5 WhoDunItDocument3 pagesActivity 5 WhoDunItRuth Danielle GasconNo ratings yet
- Bio150 Report1 Fluorescence MicrosDocument21 pagesBio150 Report1 Fluorescence MicrosRuth Danielle Gascon100% (1)
- Chem 40 April29 ReplicationDocument14 pagesChem 40 April29 ReplicationRuth Danielle GasconNo ratings yet
- Chem40 April30 Transcription and TranslationDocument32 pagesChem40 April30 Transcription and TranslationRuth Danielle GasconNo ratings yet
- Alpha Keratin: (A) Both Type I and Type II (B) The RodDocument23 pagesAlpha Keratin: (A) Both Type I and Type II (B) The RodRuth Danielle GasconNo ratings yet
- 15 April Amino AcidsDocument21 pages15 April Amino AcidsRuth Danielle GasconNo ratings yet
- 21 April EnzymesDocument22 pages21 April EnzymesRuth Danielle GasconNo ratings yet
- 29 April Lecture On Mutation Repair and DNA TechniquesDocument15 pages29 April Lecture On Mutation Repair and DNA TechniquesRuth Danielle GasconNo ratings yet
- 21 April Protein AnalysisDocument9 pages21 April Protein AnalysisRuth Danielle GasconNo ratings yet
- 14 April IntroductionDocument15 pages14 April IntroductionRuth Danielle GasconNo ratings yet
- Msds Heksana - 2 PDFDocument6 pagesMsds Heksana - 2 PDFrilmaNo ratings yet
- 10 AlgaeDocument3 pages10 AlgaeRuth Danielle GasconNo ratings yet
- 6 AcoelomatesDocument2 pages6 AcoelomatesRuth Danielle GasconNo ratings yet
- 54 3rdLEDocument1 page54 3rdLERuth Danielle GasconNo ratings yet
- 3 Bio11 Lecture Exam Notes CompilationDocument7 pages3 Bio11 Lecture Exam Notes CompilationRuth Danielle GasconNo ratings yet
- Neural TubeDocument4 pagesNeural TubeRuth Danielle GasconNo ratings yet
- Potentiometric Titration of An Acid MixtureDocument9 pagesPotentiometric Titration of An Acid MixtureRuth Danielle GasconNo ratings yet
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (400)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (121)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- ResumeDocument2 pagesResumeVisaal PradhanNo ratings yet
- Anatomy of The Psoas MuscleDocument2 pagesAnatomy of The Psoas MuscleMilos KovacevicNo ratings yet
- Material Safety Data Sheet: I. Product IdentificationDocument2 pagesMaterial Safety Data Sheet: I. Product IdentificationElMacheteDelHuesoNo ratings yet
- Yoga and Breathing: Muscles Work TogetherDocument2 pagesYoga and Breathing: Muscles Work Togethervksk1951No ratings yet
- Nephro NCP2Document2 pagesNephro NCP2hannahNo ratings yet
- Reveos BrochureDocument16 pagesReveos BrochureRushyanth KRNo ratings yet
- Elektrolit Imbalance.2013Document45 pagesElektrolit Imbalance.2013Fawzia Devi FitrianiNo ratings yet
- Immunopharmacology - IntroductionDocument51 pagesImmunopharmacology - IntroductionIsrat Jahan Surovy 2225400672No ratings yet
- 1 Cardiovascular SystemDocument6 pages1 Cardiovascular SystemWaltz PadlanNo ratings yet
- Steps To Prepare The Divine Golden BookDocument2 pagesSteps To Prepare The Divine Golden Bookdesign12No ratings yet
- Ammonium Persulfate (A3678) - Product Information SheetDocument1 pageAmmonium Persulfate (A3678) - Product Information SheetPatricio Javier Alfaro CerdaNo ratings yet
- Chlorella Spirulina DosagesDocument1 pageChlorella Spirulina DosagesYvette DawesNo ratings yet
- Anabolic Priming Posture RestoreDocument21 pagesAnabolic Priming Posture RestoreSamir KapetanovicNo ratings yet
- Vascular Surgery MedCosmos Surgery - MCQDocument34 pagesVascular Surgery MedCosmos Surgery - MCQMike GNo ratings yet
- TARIF PRODIA TAHUN 2019 Rev 2Document9 pagesTARIF PRODIA TAHUN 2019 Rev 2cici100% (1)
- Breathing For Singing-The Anatomy of RespirationDocument11 pagesBreathing For Singing-The Anatomy of RespirationAnurrag Kumar100% (3)
- Structure and Function of A Skeletal SystemDocument4 pagesStructure and Function of A Skeletal Systemapi-280073172No ratings yet
- Fluid and Electrolyte Therapy Hemant NewDocument78 pagesFluid and Electrolyte Therapy Hemant Newdrhemantparakh100% (1)
- Goodnight Beautiful-Dorothy KoomsonDocument13 pagesGoodnight Beautiful-Dorothy KoomsondarijarasetaNo ratings yet
- Music For Solo Performer by Alvin Lucier in An Investigation of Current Trends in Brainwave SonificationDocument23 pagesMusic For Solo Performer by Alvin Lucier in An Investigation of Current Trends in Brainwave SonificationJeremy WoodruffNo ratings yet
- Digestive SystemDocument5 pagesDigestive SystemFrances Anne OrbinoNo ratings yet
- Phytochemical ScreeningDocument10 pagesPhytochemical ScreeningRajesh KumarNo ratings yet
- Using The ADVAIR HFA InhalerDocument4 pagesUsing The ADVAIR HFA InhaleriveleonNo ratings yet
- AP PG Mock Part 1Document234 pagesAP PG Mock Part 1TargetNo ratings yet
- Mitosis and Meiosis Webquest 17Document3 pagesMitosis and Meiosis Webquest 17api-260817414No ratings yet
- Alveolar Bone in Health and DiseaseDocument16 pagesAlveolar Bone in Health and DiseaseVartika VermaNo ratings yet
- Artificial Neural Networks - The Tutorial With MatlabDocument23 pagesArtificial Neural Networks - The Tutorial With MatlabAndrea FieldsNo ratings yet
- Carbohydrate and Lipid Metabolism in Farm AnimalsDocument4 pagesCarbohydrate and Lipid Metabolism in Farm AnimalsPinaNo ratings yet
- Cag Views PPT Sunil PDFDocument57 pagesCag Views PPT Sunil PDFNikul J PanchalNo ratings yet
- Disease of Blood VesselsDocument5 pagesDisease of Blood VesselsKeshant SamarooNo ratings yet