You might also like
- Complete Review Lesson Plan PDFDocument8 pagesComplete Review Lesson Plan PDFapi-312176473No ratings yet
- Biopsych Trans Chap 2Document13 pagesBiopsych Trans Chap 2Chantelle SiyNo ratings yet
- Gene Therapy in Neurological DisordersFrom EverandGene Therapy in Neurological DisordersMingjie LiNo ratings yet
- 3.长征医院-MALBAC-PGD of ADPKDDocument6 pages3.长征医院-MALBAC-PGD of ADPKDzjuwindNo ratings yet
- Tema 7 Eleccion de GrupoDocument5 pagesTema 7 Eleccion de GrupoAbigail Cueva PuellesNo ratings yet
- 2021 09 14 460251v1 FullDocument22 pages2021 09 14 460251v1 Fullsyd.ashq99No ratings yet
- A Rare Case of Familial Neurogenic Diabetes Insipidu - 2021 - AACE Clinical CaseDocument4 pagesA Rare Case of Familial Neurogenic Diabetes Insipidu - 2021 - AACE Clinical CaseShuaib AhmedNo ratings yet
- 5.tang Et Al 2016Document16 pages5.tang Et Al 2016MiaNo ratings yet
- International Journal of Pharma and Bio Sciences V1 (2) 2010Document6 pagesInternational Journal of Pharma and Bio Sciences V1 (2) 2010ManikandanNo ratings yet
- Espinoza Et Al 2014Document9 pagesEspinoza Et Al 2014niltonNo ratings yet
- Risperidone Induced Seizures After Cessation of ClozapineDocument1 pageRisperidone Induced Seizures After Cessation of ClozapineAttaufiq IrawanNo ratings yet
- HDBMR - 6661860 1..7Document8 pagesHDBMR - 6661860 1..7Raul Morales VillegasNo ratings yet
- IL-7R α polymorphisms in 60 Iranian multiple sclerosis patientsDocument6 pagesIL-7R α polymorphisms in 60 Iranian multiple sclerosis patientsShukr Wesman BlbasNo ratings yet
- Doença de Batten 2004Document9 pagesDoença de Batten 2004Ana Paula BoscatoNo ratings yet
- 10 5530ijmedph 2016 3 11Document3 pages10 5530ijmedph 2016 3 11wahyu agungNo ratings yet
- UprgenyDocument9 pagesUprgenyJose Rafael Villafan BernalNo ratings yet
- Loss of P16 in Esophageal Adenocarcinoma Detected by Fluorescence in Situ Hybridization and ImmunohistochemistryDocument6 pagesLoss of P16 in Esophageal Adenocarcinoma Detected by Fluorescence in Situ Hybridization and ImmunohistochemistryTeodorNo ratings yet
- 6.fei Et Al 2015Document6 pages6.fei Et Al 2015MiaNo ratings yet
- Involvement of MnSOD Ala16Val Polymorphism in Epilepsy A Relationship With Seizure TypeDocument6 pagesInvolvement of MnSOD Ala16Val Polymorphism in Epilepsy A Relationship With Seizure TypeDetector BotsNo ratings yet
- Artículo ViroDocument13 pagesArtículo ViroNatalia BogotáNo ratings yet
- 427 EndoDocument9 pages427 EndoBioq. GallegosNo ratings yet
- National Academy of Sciences Proceedings of The National Academy of Sciences of The United States of AmericaDocument6 pagesNational Academy of Sciences Proceedings of The National Academy of Sciences of The United States of AmericamayaNo ratings yet
- Neuronal Bioenergetics and Acute Mitochondrial Dysfunction A Clue To Understanding The Central Nervous System Side Effects of Efavirenz.Document11 pagesNeuronal Bioenergetics and Acute Mitochondrial Dysfunction A Clue To Understanding The Central Nervous System Side Effects of Efavirenz.Roman_1988No ratings yet
- Gim 201618Document8 pagesGim 201618Ioana PoterasNo ratings yet
- The Landscape of Epilepsy-Related GATOR1 Variants Baldassari 2019Document11 pagesThe Landscape of Epilepsy-Related GATOR1 Variants Baldassari 2019cicasmileNo ratings yet
- Biopolym - Cell 2018 34 5 361 enDocument6 pagesBiopolym - Cell 2018 34 5 361 enАнна ШаповаловаNo ratings yet
- Biopolym - Cell 2018 34 5 361 enDocument6 pagesBiopolym - Cell 2018 34 5 361 enАнна ШаповаловаNo ratings yet
- CSF Somatic Mutations .26080Document14 pagesCSF Somatic Mutations .26080Nelson AlvarengaNo ratings yet
- Single-Nucleotide Polymorphism of The UMOD Promoter Is Associated With The Outcome of Chronic Kidney Disease PatientsDocument5 pagesSingle-Nucleotide Polymorphism of The UMOD Promoter Is Associated With The Outcome of Chronic Kidney Disease Patientssylvi293No ratings yet
- Molecular Genetic Analysis of Steroid Resistant NephroticDocument8 pagesMolecular Genetic Analysis of Steroid Resistant NephroticBeatri AyuzaNo ratings yet
- Cilia 2018Document1 pageCilia 2018Anna Beatriz Silva EspindolaNo ratings yet
- PapilomaDocument6 pagesPapilomaapi-3710742No ratings yet
- Aqac126 303Document2 pagesAqac126 303Starix UkrNo ratings yet
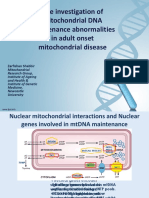
- The Investigation of Mitochondrial DNA Maintenance Abnormalities in Adult Onset Mitochondrial DiseaseDocument11 pagesThe Investigation of Mitochondrial DNA Maintenance Abnormalities in Adult Onset Mitochondrial DiseaseZarfishan ShabbirNo ratings yet
- Del Toro 2006Document5 pagesDel Toro 2006NATALIA MARTINEZ CORDOBANo ratings yet
- Infection and ImmunityDocument8 pagesInfection and Immunityivan ramaNo ratings yet
- An Infant Case of Pseudohypoaldosteronism Type1a Caused by A Novel NR3C2 VariantDocument3 pagesAn Infant Case of Pseudohypoaldosteronism Type1a Caused by A Novel NR3C2 VariantNever MindNo ratings yet
- Qi-Dan Fang Ameliorates Adriamycin-Induced Nephrotic Syndrome RatDocument9 pagesQi-Dan Fang Ameliorates Adriamycin-Induced Nephrotic Syndrome RatBeatri AyuzaNo ratings yet
- Genetic and Neurodevelopmental Spectrum of SYNGAP1-associated Intellectual Disability and EpilepsyDocument12 pagesGenetic and Neurodevelopmental Spectrum of SYNGAP1-associated Intellectual Disability and EpilepsySaya OtonashiNo ratings yet
- Epileptic Encephalopathies SYNGAP1Document7 pagesEpileptic Encephalopathies SYNGAP1Saya OtonashiNo ratings yet
- Knockdown of NUPR1 Inhibits The Proliferation of Glioblastoma Cells Via ERK1 - 2, p38 MAPK and Caspase-3Document12 pagesKnockdown of NUPR1 Inhibits The Proliferation of Glioblastoma Cells Via ERK1 - 2, p38 MAPK and Caspase-3郭耿良No ratings yet
- Department of Veterinary Biosciences, College of Veterinary Medicine, The Ohio State University, Columbus, Ohio 43210-1093Document9 pagesDepartment of Veterinary Biosciences, College of Veterinary Medicine, The Ohio State University, Columbus, Ohio 43210-1093FrontiersNo ratings yet
- Molecular Diagnosis ofDocument67 pagesMolecular Diagnosis ofNadir KoçakNo ratings yet
- Increased Expression of Heparanase in Puromycin Aminonucleoside NephrosisDocument10 pagesIncreased Expression of Heparanase in Puromycin Aminonucleoside NephrosisFauziana NurhanisahNo ratings yet
- Liu 2005Document7 pagesLiu 2005Javier Cerda InfanteNo ratings yet
- Digenic Parkinsonism: Investigation of The Synergistic Effects of PRKN and LRRK2Document5 pagesDigenic Parkinsonism: Investigation of The Synergistic Effects of PRKN and LRRK2SakinaNo ratings yet
- Jack 2Document5 pagesJack 2Josue BarralNo ratings yet
- 2019 Article 421DTNDocument8 pages2019 Article 421DTNDulce Maria AngelesNo ratings yet
- Expanded Phenotypes and Outcomes Among 256 Lgi1/Caspr2-Igg-Positive PatientsDocument14 pagesExpanded Phenotypes and Outcomes Among 256 Lgi1/Caspr2-Igg-Positive PatientsaaboffNo ratings yet
- Expression of The Nociceptin Precursor and NocicepDocument7 pagesExpression of The Nociceptin Precursor and NocicepRENAULTNo ratings yet
- Sustained Delivery and Expression of DNA Encapsulated in Polymeric NanoparticlesDocument10 pagesSustained Delivery and Expression of DNA Encapsulated in Polymeric NanoparticlesAngy ÑacatoNo ratings yet
- Migdalska-Richards Et Al-2016-Journal of NeurochemistryDocument14 pagesMigdalska-Richards Et Al-2016-Journal of Neurochemistryjazz castañedaNo ratings yet
- Loss in PKC Epsilon Causes Downregulation of MnSOD and BDNF Expression in Neurons of Alzheimer's Disease HippocampusDocument17 pagesLoss in PKC Epsilon Causes Downregulation of MnSOD and BDNF Expression in Neurons of Alzheimer's Disease HippocampusAnonymous mt8XmkRWNo ratings yet
- A 5 Year Old Boy With Super Refractory Status Epilepticus and RANBP2 Variant WarrantingDocument8 pagesA 5 Year Old Boy With Super Refractory Status Epilepticus and RANBP2 Variant WarrantingAlice MaulisovaNo ratings yet
- Url-2003 Parvovirus en Neuronas de GatoDocument5 pagesUrl-2003 Parvovirus en Neuronas de GatocristianNo ratings yet
- Pluripotent Anti-Inflammatory Immunomodulatory Effects of Papaverine Against Cerebral Ischemic-Reperfusion InjuryDocument23 pagesPluripotent Anti-Inflammatory Immunomodulatory Effects of Papaverine Against Cerebral Ischemic-Reperfusion InjuryDragomir MirunaNo ratings yet
- Maed 07 271Document6 pagesMaed 07 271MARDHIANo ratings yet
- 10.0000@Www - Alliedacademies.org@nod2card15 Rs2066845 Polymorphism in Children With Acute AppendicitisDocument6 pages10.0000@Www - Alliedacademies.org@nod2card15 Rs2066845 Polymorphism in Children With Acute Appendicitisefancoolhand09No ratings yet
- Neuropharmacology: Sciverse SciencedirectDocument8 pagesNeuropharmacology: Sciverse SciencedirectShawnNo ratings yet
- Ni Hms 614459Document20 pagesNi Hms 614459Prakash GudsoorkarNo ratings yet
- Bok:978 81 322 0958 4 PDFDocument304 pagesBok:978 81 322 0958 4 PDFSamiNo ratings yet
- Science and Technology EssayDocument18 pagesScience and Technology EssayDean Kurt SaroopNo ratings yet
- 1 Examen 2 Trimestre BiologiaDocument3 pages1 Examen 2 Trimestre Biologianacho urbina rodriguezNo ratings yet
- Biology A100: Principles of Biology Tentative Syllabus: Clew7@occ - Cccd.eduDocument10 pagesBiology A100: Principles of Biology Tentative Syllabus: Clew7@occ - Cccd.eduDuy NguyenNo ratings yet
- Ethical Issues Surrounding Gene TherapyDocument4 pagesEthical Issues Surrounding Gene TherapyChristianne CapuaNo ratings yet
- Model Questions: Unit - IDocument8 pagesModel Questions: Unit - IRavikanthNo ratings yet
- Test Bank MetabolismDocument6 pagesTest Bank MetabolismWanga CharleneNo ratings yet
- Anti Bastos LawDocument4 pagesAnti Bastos Lawchandy RendajeNo ratings yet
- K Biotech 123Document2 pagesK Biotech 123Dave DhruvNo ratings yet
- Duration: 180 Mins: © Ocr 2020. You May Photocopy This Page. 1 of 30 Created in ExambuilderDocument30 pagesDuration: 180 Mins: © Ocr 2020. You May Photocopy This Page. 1 of 30 Created in ExambuilderIfeanyi AdemijuNo ratings yet
- Fundamentals of GeneticsDocument16 pagesFundamentals of GeneticsshriNo ratings yet
- Essential Molecular BiologyDocument19 pagesEssential Molecular BiologySandra JelynNo ratings yet
- pRS415Document12 pagespRS415DiegoNo ratings yet
- Human Genome ProjectDocument42 pagesHuman Genome ProjectGopikagopinathan RajeshNo ratings yet
- Human ORFeome Gateway Entry Vector PENTR223Document6 pagesHuman ORFeome Gateway Entry Vector PENTR223Dave RapaconNo ratings yet
- An TR Hop Ology 101 FinalDocument224 pagesAn TR Hop Ology 101 FinalJunaid AliNo ratings yet
- The Investigation of Mitochondrial DNA Maintenance Abnormalities in Adult Onset Mitochondrial DiseaseDocument11 pagesThe Investigation of Mitochondrial DNA Maintenance Abnormalities in Adult Onset Mitochondrial DiseaseZarfishan ShabbirNo ratings yet
- Principles of Population GeneticsDocument7 pagesPrinciples of Population GeneticsGiulia Magri RibeiroNo ratings yet
- Ayala - Teleological Explanations in Evolutionary BiologyDocument16 pagesAyala - Teleological Explanations in Evolutionary BiologyRodrigo CarvalhoNo ratings yet
- Jurnal Buah Naga PDFDocument7 pagesJurnal Buah Naga PDFAmelia GustinNo ratings yet
- 57-1-2 BiologyDocument15 pages57-1-2 BiologyRavneet KaurNo ratings yet
- Prep3 Final Revision..Document26 pagesPrep3 Final Revision..Amira Hekal0% (1)
- Transcription Factor-Based Biosensors in Biotechnology - CurrentDocument12 pagesTranscription Factor-Based Biosensors in Biotechnology - CurrentConcetta SantiniNo ratings yet
- Chapter 16 - Origin and Evolution of Microbial Life - PPTX DimaporoDocument39 pagesChapter 16 - Origin and Evolution of Microbial Life - PPTX DimaporoRamon Chito S. PudelananNo ratings yet
- GREDocument7 pagesGREBoris ShalomovNo ratings yet
- Learning Objectives: Module 1 - Physical SelfDocument4 pagesLearning Objectives: Module 1 - Physical SelfMiss MegzzNo ratings yet
- Interaction of GenesDocument15 pagesInteraction of GenesAmit SharmaNo ratings yet
- Medical Biochemistry 6E Sep 21 2022 John W Baynes Full ChapterDocument67 pagesMedical Biochemistry 6E Sep 21 2022 John W Baynes Full Chaptermichael.yung118100% (3)