You might also like
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Patanjali Yoga Sutras by Sri Sri Ravi Shankar PDFDocument6 pagesPatanjali Yoga Sutras by Sri Sri Ravi Shankar PDFAnil Mehta33% (3)
- Total Number of Posts Advertised 575Document11 pagesTotal Number of Posts Advertised 575Anil MehtaNo ratings yet
- Death of Salesman As TragedyDocument12 pagesDeath of Salesman As TragedyAnil Mehta100% (2)
- KAS Notification 2011Document4 pagesKAS Notification 2011Anil MehtaNo ratings yet
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5795)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (345)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (400)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (121)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- Cell Biology of Plant Nematode ParasitismDocument277 pagesCell Biology of Plant Nematode ParasitismkamalNo ratings yet
- NematodeschapterDocument9 pagesNematodeschapterArjelyn Mae D BuendiaNo ratings yet
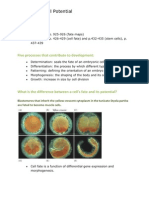
- Cell Fate and Cell PotentialDocument12 pagesCell Fate and Cell PotentialSavannah Simone PetrachenkoNo ratings yet
- Development of The Nervous System 3rd Edition Sanes Test BankDocument35 pagesDevelopment of The Nervous System 3rd Edition Sanes Test Bankbateddivisor51zwed100% (20)
- Journal Pre-Proofs: Food ChemistryDocument34 pagesJournal Pre-Proofs: Food ChemistryJosueChalloNo ratings yet
- Nematode BehaviourDocument445 pagesNematode Behaviourdouglasman100% (4)
- Chinise MedicineDocument13 pagesChinise Medicinedyg dygNo ratings yet
- Caenorhabditis Elegans A Pluricellular Model - 2010 - Biochimica Et BiophysicaDocument9 pagesCaenorhabditis Elegans A Pluricellular Model - 2010 - Biochimica Et BiophysicaDesmond BohamNo ratings yet
- Proteomic Analysis of Caenorhabditis Elegans Wound Model Reveals Novel Molecular Players Involved in RepairDocument15 pagesProteomic Analysis of Caenorhabditis Elegans Wound Model Reveals Novel Molecular Players Involved in RepairalessioNo ratings yet
- Microbial Biotechnology - 2021 - Balaguer - Lipoteichoic Acid From Bifidobacterium Animalis Subsp Lactis BPL1 A NovelDocument12 pagesMicrobial Biotechnology - 2021 - Balaguer - Lipoteichoic Acid From Bifidobacterium Animalis Subsp Lactis BPL1 A NovelStephanie M de LeonNo ratings yet
- Anti AgingDocument6 pagesAnti AgingZubair Akram JanjuaNo ratings yet
- ToxocaraDocument23 pagesToxocaraDaniela Hernandez SalazarNo ratings yet
- Gerobiotics Probiotics Targeting Fundamental AgingDocument35 pagesGerobiotics Probiotics Targeting Fundamental AgingAnkshu RayNo ratings yet
- Wormbot PaperDocument13 pagesWormbot Paperapi-545982271No ratings yet
- The Development of FrogDocument13 pagesThe Development of FrogRhonnel Manatad AlburoNo ratings yet
- W.P. de Voogt Eds. Reviews of Environmental Contamination and Toxicology Volume 237Document204 pagesW.P. de Voogt Eds. Reviews of Environmental Contamination and Toxicology Volume 237Gabrielle Abreu NunesNo ratings yet
- Nucleotide Sugar TransportersDocument8 pagesNucleotide Sugar TransportersCosmin BarbosNo ratings yet
- RNAi Feeding TimmonsDocument10 pagesRNAi Feeding TimmonsfrostyNo ratings yet
- Longevity Trends 2020Document51 pagesLongevity Trends 2020Fabiana SouzaNo ratings yet
- (Handbook of Zoology) Andreas Schmidt-rhaesa-Nematoda-De Gruyter (2013)Document777 pages(Handbook of Zoology) Andreas Schmidt-rhaesa-Nematoda-De Gruyter (2013)Jefri Amfotis100% (1)
- Danger Avoidance Can Be Genetically Encoded For Four Generations, Say Biologists - Scientists Found That Learned Behaviors in CDocument3 pagesDanger Avoidance Can Be Genetically Encoded For Four Generations, Say Biologists - Scientists Found That Learned Behaviors in Cdamdidam1No ratings yet
- Practical Manual of Developmental BiologyDocument93 pagesPractical Manual of Developmental Biologyarushi4702No ratings yet
- C. Elegans Genetic PortaitDocument26 pagesC. Elegans Genetic PortaitbelovedaffectionNo ratings yet
- Anthelmintic DrugsDocument13 pagesAnthelmintic DrugsMaria Grazia SantoroNo ratings yet
- 2009-High Resolution Map of Caenorhabditis Elegans Gap Junction ProteinsDocument15 pages2009-High Resolution Map of Caenorhabditis Elegans Gap Junction ProteinsyicinenNo ratings yet
- Apa Short Essay FormatDocument6 pagesApa Short Essay Formatwwpadwwhd100% (2)
- Nematodes As Environmental IndicatorsDocument341 pagesNematodes As Environmental Indicatorsgupta_pun100% (6)
- Developmental GeneticsDocument20 pagesDevelopmental GeneticsabhishekNo ratings yet
- Final Capstone Poster-Mann PDFDocument1 pageFinal Capstone Poster-Mann PDFAmy Nottingham-MartinNo ratings yet
- Anders Olsen, Matthew S. Gill Eds. Ageing Lessons From C. Elegans PDFDocument433 pagesAnders Olsen, Matthew S. Gill Eds. Ageing Lessons From C. Elegans PDFPhil BoileauNo ratings yet