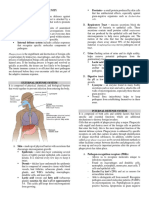
You might also like
- Blotting TechniqueDocument34 pagesBlotting TechniqueUmair100% (1)
- What is blotting techniquesDocument28 pagesWhat is blotting techniquesShivraj JadhavNo ratings yet
- Prepared By: Verna Jean M. Magdayao 3/Bsbiology/ADocument56 pagesPrepared By: Verna Jean M. Magdayao 3/Bsbiology/AKathleya PeñaNo ratings yet
- RDT Hybridization Method DetectionDocument13 pagesRDT Hybridization Method DetectionAnanya SinghNo ratings yet
- Blotting TechniquesDocument28 pagesBlotting TechniquesJoshua MuegoNo ratings yet
- Unit 1: Structural GenomicsDocument4 pagesUnit 1: Structural GenomicsLavanya ReddyNo ratings yet
- Western BlotDocument40 pagesWestern BlotЂорђе100% (1)
- cDNA Libraries and Gene CloningDocument8 pagescDNA Libraries and Gene CloningRoberto RomeroNo ratings yet
- "In Situ" Hybridization TechniquesDocument41 pages"In Situ" Hybridization TechniquesNT RavindranNo ratings yet
- Southern Northern and Western BlottingDocument48 pagesSouthern Northern and Western BlottingharithagenNo ratings yet
- Crispr Cas HajarDocument21 pagesCrispr Cas HajarHajira Fatima100% (1)
- Bacterial GeneticsDocument9 pagesBacterial GeneticsExamville.comNo ratings yet
- Purification of DNADocument14 pagesPurification of DNAalivetutorsNo ratings yet
- DNA Extraction Whole BloodDocument27 pagesDNA Extraction Whole BloodW0% (1)
- Blotting TechniquesDocument3 pagesBlotting TechniquesabidaiqbalansariNo ratings yet
- Restriction Enzymes PDFDocument7 pagesRestriction Enzymes PDFmanoj_rkl_07No ratings yet
- Detection of Nucleic AcidDocument13 pagesDetection of Nucleic AcidAfrah AlatasNo ratings yet
- Types of Electrophoresis and DNA Fingerprinting TechniquesDocument73 pagesTypes of Electrophoresis and DNA Fingerprinting TechniquesJelsea AmarradorNo ratings yet
- Bi0505 LabDocument102 pagesBi0505 LabJatinMittalNo ratings yet
- PCR Types and Its ApplicationsDocument68 pagesPCR Types and Its ApplicationsShefali Pawar100% (1)
- Selection of Recombinant ClonesDocument2 pagesSelection of Recombinant ClonesDeepika KVNo ratings yet
- PCR, RT-PCR, Nested-Pcr, Multiplex PCR, Quantitative PCRDocument116 pagesPCR, RT-PCR, Nested-Pcr, Multiplex PCR, Quantitative PCRYunizardiNo ratings yet
- Cloning VectorsDocument8 pagesCloning Vectorssanchit100% (3)
- microRNA, Gene Mapping and Gene SequencingDocument33 pagesmicroRNA, Gene Mapping and Gene SequencingAdarsh ShrivastavaNo ratings yet
- Protein Microarray Naveed Up MushtaqDocument16 pagesProtein Microarray Naveed Up MushtaqAngumaniNo ratings yet
- Extraction of Genomic DNA: G.Umamaheswaran PH.D Scholar JipmerDocument24 pagesExtraction of Genomic DNA: G.Umamaheswaran PH.D Scholar Jipmerpharmaguy111No ratings yet
- Southern Blotting Techniques ExplainedDocument50 pagesSouthern Blotting Techniques ExplainedSandeep Otari100% (1)
- VectorsDocument6 pagesVectorsAssad MustafaNo ratings yet
- Innate Immunity Chapter SummaryDocument11 pagesInnate Immunity Chapter SummaryOrhan AsdfghjklNo ratings yet
- Dna RepairDocument20 pagesDna RepairEaron Van JaboliNo ratings yet
- Recombinant DNA Technology: Dr. P. Balaji Head in Biotechnology MGR College, HosurDocument78 pagesRecombinant DNA Technology: Dr. P. Balaji Head in Biotechnology MGR College, HosurBalaji Paulraj100% (1)
- Linkage: Harshraj Subhash Shinde KKW, Cabt, NashikDocument14 pagesLinkage: Harshraj Subhash Shinde KKW, Cabt, Nashiksivaram888No ratings yet
- Biotecnology and Its ApplicationDocument87 pagesBiotecnology and Its ApplicationRangaswamyBiligiraiah100% (1)
- Electrophoretic Mobility Shift Assay - Wikipedia, The Free EncyclopediaDocument3 pagesElectrophoretic Mobility Shift Assay - Wikipedia, The Free EncyclopediaShailendra YadavNo ratings yet
- Cot CurveDocument16 pagesCot CurveVidyasagar Deshpande100% (1)
- Real-Time PCR Applications - Presentation by Nasr SinjilawiDocument69 pagesReal-Time PCR Applications - Presentation by Nasr SinjilawiMolecular_Diagnostics_KKUHNo ratings yet
- Practical 1 Lab Safety and DNA Isolation PDFDocument14 pagesPractical 1 Lab Safety and DNA Isolation PDFAbery AuNo ratings yet
- Practical Hand Outs-Molecular Biology (BIO302)Document33 pagesPractical Hand Outs-Molecular Biology (BIO302)Anaya MalikNo ratings yet
- Inheritance Biology EDUNCLEDocument54 pagesInheritance Biology EDUNCLEEvita Almeida100% (1)
- Plasmid VectorsDocument31 pagesPlasmid VectorsDebajeet GhoshNo ratings yet
- Proteomics IntroductionDocument39 pagesProteomics Introductionnariel67% (3)
- Dna Sequencing: DR Z Chikwambi BiotechnologyDocument110 pagesDna Sequencing: DR Z Chikwambi BiotechnologyMutsawashe Munetsi100% (1)
- Sequecing - Khan AcademyDocument16 pagesSequecing - Khan AcademyMaryem SafdarNo ratings yet
- DNA DamageDocument3 pagesDNA DamageAnonymous yNWMB80ffFNo ratings yet
- Sequencing TechnologiesDocument25 pagesSequencing TechnologiesOhhh OkayNo ratings yet
- Microbiology Methods of Monitoring PopulationsDocument17 pagesMicrobiology Methods of Monitoring PopulationsStephen MooreNo ratings yet
- Methods To Study Soil Microbial DiversityDocument25 pagesMethods To Study Soil Microbial Diversityrd2165scribdNo ratings yet
- NanomedicineDocument64 pagesNanomedicinePrabisha PrabhakaranNo ratings yet
- ChromosomesDocument4 pagesChromosomesعامر جدونNo ratings yet
- A Plasmid Is A Small DNA Molecule Within A Cell That Is Physically Separated From A Chromosomal DNA and Can Replicate IndependentlyDocument5 pagesA Plasmid Is A Small DNA Molecule Within A Cell That Is Physically Separated From A Chromosomal DNA and Can Replicate Independentlyyaqoob008No ratings yet
- Single-Nucleotide PolymorphismDocument21 pagesSingle-Nucleotide PolymorphismdearbhupiNo ratings yet
- DNA sequencing: Sanger methodDocument14 pagesDNA sequencing: Sanger methodummu sahdhiyya3No ratings yet
- Molecular MarkersDocument42 pagesMolecular Markerssaroj Burlakoti100% (1)
- CELL CULTURE LINESDocument31 pagesCELL CULTURE LINESRamesh BeniwalNo ratings yet
- DNA Manipulative EnzymesDocument17 pagesDNA Manipulative EnzymesZain Ul AbedienNo ratings yet
- Guide To The Classification and Identification of The Actinomycetes and Their Antibiotics by Waksman, Selman A. (Selman Abraham), 1888-1973 Lechevalier, Hubert ADocument264 pagesGuide To The Classification and Identification of The Actinomycetes and Their Antibiotics by Waksman, Selman A. (Selman Abraham), 1888-1973 Lechevalier, Hubert AVictor R SatrianiNo ratings yet
- Gene MappingDocument4 pagesGene MappingOlib OlieNo ratings yet
- ABT 227 - Course Outline Introduction To Molecular Biology (3335)Document1 pageABT 227 - Course Outline Introduction To Molecular Biology (3335)justevansiNo ratings yet
- Monoclonal Antibodies Against Bacteria: Volume IIIFrom EverandMonoclonal Antibodies Against Bacteria: Volume IIIAlberto J. L. MacarioNo ratings yet
- Img20211212 16211914Document1 pageImg20211212 16211914Dayananda SalamNo ratings yet
- Ukhrul AtRDocument8 pagesUkhrul AtRDayananda SalamNo ratings yet
- Bishnupur AtRDocument5 pagesBishnupur AtRDayananda SalamNo ratings yet
- II. Statement Showing Progressive Lifting of Foodgrains Under Various Schemes During The Period Starting From 1st October, 2021Document1 pageII. Statement Showing Progressive Lifting of Foodgrains Under Various Schemes During The Period Starting From 1st October, 2021Dayananda SalamNo ratings yet
- 5 6264955940114531395Document49 pages5 6264955940114531395anuragNo ratings yet
- Short Term: Availability of Platforrm Scales in The DepotDocument6 pagesShort Term: Availability of Platforrm Scales in The DepotDayananda SalamNo ratings yet
- Swacch Bharat Abhiyan 2.0Document2 pagesSwacch Bharat Abhiyan 2.0Dayananda SalamNo ratings yet
- Tumor Suppressor GeneDocument21 pagesTumor Suppressor GeneDayananda SalamNo ratings yet
- Biosynthesis - Wikipedia, The Free EncyclopediaDocument15 pagesBiosynthesis - Wikipedia, The Free EncyclopediaDayananda SalamNo ratings yet
- Infection of Cells by BacteriaDocument8 pagesInfection of Cells by BacteriaDayananda SalamNo ratings yet
- LifeA June2016Document44 pagesLifeA June2016Dev ShuklaNo ratings yet
- Bioinert HPLCDocument8 pagesBioinert HPLCDayananda SalamNo ratings yet
- Bacterial Adhesion and EntryDocument13 pagesBacterial Adhesion and EntryZeus SaldanaNo ratings yet
- Marker Assisted SelectionDocument6 pagesMarker Assisted SelectionDayananda SalamNo ratings yet
- User Manual: Avira Antivir PremiumDocument99 pagesUser Manual: Avira Antivir PremiumDayananda SalamNo ratings yet
- Chargeswitch Direct 8-Well or 96 Gdna Kits: Catalog Nos. Cs11205, Cs11206, Cs11209Document4 pagesChargeswitch Direct 8-Well or 96 Gdna Kits: Catalog Nos. Cs11205, Cs11206, Cs11209Dayananda SalamNo ratings yet
- Northern Blotting Process & Technique for Detecting Gene ExpressionDocument18 pagesNorthern Blotting Process & Technique for Detecting Gene ExpressionAbdul Ghani100% (1)
- Alexis Bishop - PCR Lab ReportDocument2 pagesAlexis Bishop - PCR Lab Reportapi-353843842No ratings yet
- Immunological Techniques IDocument28 pagesImmunological Techniques Imartinian_4everNo ratings yet
- Real-Time PCR Applications - Presentation by Nasr SinjilawiDocument69 pagesReal-Time PCR Applications - Presentation by Nasr SinjilawiMolecular_Diagnostics_KKUHNo ratings yet
- Bio PresentationDocument2 pagesBio Presentationchrisshit100% (1)
- Blotting Assignment by MPhil 5,12Document31 pagesBlotting Assignment by MPhil 5,12Amina ChNo ratings yet
- Agarose Vs PolyacrylamideDocument1 pageAgarose Vs PolyacrylamideJadie WellingNo ratings yet
- DNA Extraction Lab ReportDocument7 pagesDNA Extraction Lab ReportNazurah IbrahimNo ratings yet
- Specifikacio Altalanos Vegyszer - 1. - NeurologiaDocument3 pagesSpecifikacio Altalanos Vegyszer - 1. - Neurologiabotond77No ratings yet
- Gel Electrophoresis Basics Worksheet: NameDocument2 pagesGel Electrophoresis Basics Worksheet: NameJosel SoteroNo ratings yet
- BSc Student Presents Agarose Gel ElectrophoresisDocument18 pagesBSc Student Presents Agarose Gel ElectrophoresisSathvik BangiramaneNo ratings yet
- DNA Extraction and Gel ElectrophoresisDocument31 pagesDNA Extraction and Gel Electrophoresisanimathz0% (1)
- Protein Lab ReportDocument7 pagesProtein Lab ReportFariha0% (1)
- Lab 5 Gel ElectrophoresisDocument2 pagesLab 5 Gel Electrophoresishew2215No ratings yet
- K26. Aspek Fisika Dalam BiomolekulDocument68 pagesK26. Aspek Fisika Dalam BiomolekulFalih KepahiangNo ratings yet
- DGGE Analysis of Symbiodinium ITS2 for Strain DifferentiationDocument9 pagesDGGE Analysis of Symbiodinium ITS2 for Strain Differentiationsathish.botNo ratings yet
- Immunoelectrophoresis: Ankur Gautam, PHD., Merck High End Skill Development Centre Chandigarh, India 20 April, 2020Document16 pagesImmunoelectrophoresis: Ankur Gautam, PHD., Merck High End Skill Development Centre Chandigarh, India 20 April, 2020Anjali SinghNo ratings yet
- Biosensor Lab (SBFT) : Standard Operating Procedure FOR Gel Electrophoresis UnitDocument2 pagesBiosensor Lab (SBFT) : Standard Operating Procedure FOR Gel Electrophoresis UnitVijayDubeyNo ratings yet
- Agarose Gel ElectrophoresisDocument4 pagesAgarose Gel ElectrophoresisTarequl Islam NishadNo ratings yet
- Fermentas Coa Sm0671Document3 pagesFermentas Coa Sm0671Nguyễn Hoài ThuNo ratings yet
- Sds Page ProtocolDocument2 pagesSds Page ProtocolswapnilNo ratings yet
- FRAP Tutorial EMBO DebrecenDocument55 pagesFRAP Tutorial EMBO DebrecenYunonNo ratings yet
- 9 DNA TroubleshootingDocument6 pages9 DNA TroubleshootingRicha AroraNo ratings yet
- Detect Protein-DNA Binding with EMSADocument19 pagesDetect Protein-DNA Binding with EMSAAll rounders studyNo ratings yet
- Gel Electrophoresis - Wikipedia, The Free EncyclopediaDocument10 pagesGel Electrophoresis - Wikipedia, The Free EncyclopediaShailendra YadavNo ratings yet
- CE KitDocument12 pagesCE KitJohn Tyler KaufmanNo ratings yet
- TR0051 Elute From PolyacrylamideDocument2 pagesTR0051 Elute From PolyacrylamideKasagalaNo ratings yet
- Principles of Gel ElectrophoresisDocument6 pagesPrinciples of Gel ElectrophoresisCarmen Lopez100% (1)
- CSB E13415hDocument14 pagesCSB E13415hhuripNo ratings yet
- Prestained Protein Ladder for SDS-PAGE & Western BlottingDocument2 pagesPrestained Protein Ladder for SDS-PAGE & Western BlottingTran TuanNo ratings yet