You might also like
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5795)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (400)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (345)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (121)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- (Methods in Enzymology 529) Jon Lorsch (Eds.) - Laboratory Methods in Enzymology - DNA-Academic Press (2013)Document379 pages(Methods in Enzymology 529) Jon Lorsch (Eds.) - Laboratory Methods in Enzymology - DNA-Academic Press (2013)Gabriela QueirozNo ratings yet
- Muscular SystemDocument8 pagesMuscular Systemtheodore_estradaNo ratings yet
- Physical Aging Process: When Does Old Age Begin?Document32 pagesPhysical Aging Process: When Does Old Age Begin?ZeusKimNo ratings yet
- 7396 26182 1 PBDocument17 pages7396 26182 1 PBdavideNo ratings yet
- Robbins Ch. 27 Peripheral Nerve and Skeletal Muscle Review QuestionsDocument2 pagesRobbins Ch. 27 Peripheral Nerve and Skeletal Muscle Review QuestionsPA2014100% (1)
- Pancytopenia 170119201048 PDFDocument33 pagesPancytopenia 170119201048 PDFDabogski France100% (1)
- Dermatosis Del Embarazo 2008Document8 pagesDermatosis Del Embarazo 2008rizqi_cepiNo ratings yet
- Human Color VisionDocument368 pagesHuman Color VisionAriel CarvalhoNo ratings yet
- Protein Synthesis: Transcription & TranslationDocument4 pagesProtein Synthesis: Transcription & TranslationtshahriyarNo ratings yet
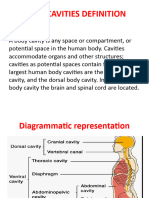
- Body Cavities PPT & Anatomical SystemDocument10 pagesBody Cavities PPT & Anatomical Systemnandhini raguNo ratings yet
- Chap 3 Paper - Theoretical Frameworks and Nursing ResearchDocument9 pagesChap 3 Paper - Theoretical Frameworks and Nursing ResearchEtty EriyantiNo ratings yet
- PCRDocument31 pagesPCRsrgNo ratings yet
- Owen A O'Connor, Stephen M Ansell, John F Seymo 230518 053705Document514 pagesOwen A O'Connor, Stephen M Ansell, John F Seymo 230518 053705Juan RivasNo ratings yet
- Abnormal Cervical CytologyDocument21 pagesAbnormal Cervical CytologyNatalia HaikaliNo ratings yet
- ProteusDocument20 pagesProteusMonishaNo ratings yet
- Gavrila Catalin 1Document1 pageGavrila Catalin 1Doina RusuNo ratings yet
- Inflammatory Bowel DiseaseDocument27 pagesInflammatory Bowel DiseaseMihai VladescuNo ratings yet
- Biology Paper 2 SLDocument16 pagesBiology Paper 2 SLCandela HerreraNo ratings yet
- Patel Hospital: Molecular PathologyDocument1 pagePatel Hospital: Molecular PathologyFarrukh NaveedNo ratings yet
- Mercy KillingDocument37 pagesMercy KillingRamesh Babu Tatapudi100% (1)
- Laporan Asuhan Keperawatan Bahasa InggrisDocument8 pagesLaporan Asuhan Keperawatan Bahasa InggrisBagus Wijanarko100% (1)
- Lypm Immunity SystemDocument22 pagesLypm Immunity SystemPhoenix JudgeNo ratings yet
- University of Cambridge International Examinations General Certificate of Education Ordinary LevelDocument12 pagesUniversity of Cambridge International Examinations General Certificate of Education Ordinary Levelmstudy123456No ratings yet
- Approach To Skin Adnexal TumorsDocument20 pagesApproach To Skin Adnexal TumorsNirmalya Chakrabarti100% (1)
- Microbial Interactions With The Host in Periodontal DiseasesDocument74 pagesMicrobial Interactions With The Host in Periodontal Diseasesrupali100% (2)
- Sturge Weber Syndrome Case Report StudyDocument11 pagesSturge Weber Syndrome Case Report StudyOphtalmology100% (3)
- Munroejacquelinedecember 5 Edps 651 FinalexamDocument22 pagesMunroejacquelinedecember 5 Edps 651 Finalexamapi-259390419No ratings yet
- Nyas12203Becoming Conscious The Science of MindfulnessDocument18 pagesNyas12203Becoming Conscious The Science of MindfulnesscgrossNo ratings yet
- Jam 14704Document17 pagesJam 14704Raja ChandraNo ratings yet
- Thesis On Retinitis PigmentosaDocument7 pagesThesis On Retinitis Pigmentosalbbzfoxff100% (2)