You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (345)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (121)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (400)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Case Study For DMDocument5 pagesCase Study For DMJermeLou Bao100% (1)
- Circulatory System WorksheetDocument2 pagesCirculatory System Worksheetrubixcube4550% (12)
- DR - Iniesta ReportDocument97 pagesDR - Iniesta ReportRaceRadioNo ratings yet
- Presentation of Audrey M Battu From Clinton Health Access Initiative (CHAI)Document17 pagesPresentation of Audrey M Battu From Clinton Health Access Initiative (CHAI)bobbyramakant0% (1)
- Home Visit Steps: Perform The Bag TechniqueDocument4 pagesHome Visit Steps: Perform The Bag TechniqueRasec Nayr CoseNo ratings yet
- Pengaruh Kompresi RJP Dengan Metronom Terhadap Kualitas Kompresi RJP Yang Dilakukan Mahasiswa PerawatDocument7 pagesPengaruh Kompresi RJP Dengan Metronom Terhadap Kualitas Kompresi RJP Yang Dilakukan Mahasiswa PerawatGabriela Adeleda LeiwakabessyNo ratings yet
- 2011-2012 Annual Report Department of Pediatrics Saint Louis University School of MedicineDocument131 pages2011-2012 Annual Report Department of Pediatrics Saint Louis University School of Medicineslusompeds0% (1)
- Nurses Notes TBDocument4 pagesNurses Notes TBSanvar Mal Soni0% (2)
- Ataxia 10Document19 pagesAtaxia 10Cristina GhizdavetNo ratings yet
- PROGRAMME ABSTRACT BOOK IACS ES 4 7 Oct 2023 Timisoara Romania - CORRDocument164 pagesPROGRAMME ABSTRACT BOOK IACS ES 4 7 Oct 2023 Timisoara Romania - CORRAlex MircescuNo ratings yet
- Complete Protein Antigens and Low - Molecular-Weight Substances - Complete Protein Antigens, Such As AnimalDocument1 pageComplete Protein Antigens and Low - Molecular-Weight Substances - Complete Protein Antigens, Such As AnimalApple BananaNo ratings yet
- Psychiatry 101Document17 pagesPsychiatry 101Vaso Talakvadze0% (1)
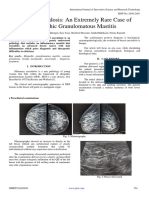
- Breast Sarcoidosis An Extremely Rare Case of Idiopathic Granulomatous MastitisDocument3 pagesBreast Sarcoidosis An Extremely Rare Case of Idiopathic Granulomatous MastitisInternational Journal of Innovative Science and Research TechnologyNo ratings yet
- UNIT 4 and UNIT 5 ISDocument9 pagesUNIT 4 and UNIT 5 ISMarinelle TumanguilNo ratings yet
- Case Scenario/Situation Answer/Explanation: NCM 101 RLE - Health AssessmentDocument2 pagesCase Scenario/Situation Answer/Explanation: NCM 101 RLE - Health AssessmentmaryNo ratings yet
- The Body Nutrition PortionDocument35 pagesThe Body Nutrition PortionHerditya putri rahmaNo ratings yet
- TMJ SlidesDocument113 pagesTMJ SlidesRah Ma GhassanNo ratings yet
- Oxygenation Structure of The Respiratory System: Pulmonary VentilationDocument40 pagesOxygenation Structure of The Respiratory System: Pulmonary VentilationCaitlin G.No ratings yet
- The LA Classification of GERDDocument2 pagesThe LA Classification of GERDelisa100% (1)
- Step2ck Practice TestDocument67 pagesStep2ck Practice TestAlf CarlNo ratings yet
- The Revised National Tuberculosis Control ProgrammeDocument2 pagesThe Revised National Tuberculosis Control ProgrammesuryaslideNo ratings yet
- Smoking Poor Dietary Habits Sedentary Lifestyle: Lung Cancer Diabetes Heart DiseaseDocument2 pagesSmoking Poor Dietary Habits Sedentary Lifestyle: Lung Cancer Diabetes Heart DiseaseKien Dela CruzNo ratings yet
- Diarrhea in ChildrenDocument42 pagesDiarrhea in ChildrenIPNATC NEPALNo ratings yet
- Addendum Re ExpertsDocument11 pagesAddendum Re ExpertsrpclarkeNo ratings yet
- PWAT (Panographic Wound Assesment Tool) RevisedDocument4 pagesPWAT (Panographic Wound Assesment Tool) RevisedYunie ArmyatiNo ratings yet
- First Aid Kits & Their ImportanceDocument25 pagesFirst Aid Kits & Their ImportanceJennette BelliotNo ratings yet
- Bhopal (M.P.) : Assignment OnDocument9 pagesBhopal (M.P.) : Assignment OnamitNo ratings yet
- UNIT 12" It Really WorksDocument2 pagesUNIT 12" It Really WorksZulfa raniNo ratings yet
- Food HandlersDocument27 pagesFood HandlersNickz AgbayaniNo ratings yet
- E Stimation of HemoglobinDocument13 pagesE Stimation of HemoglobinSANANo ratings yet