You might also like
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (400)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (121)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- Hawassa University Daye Campus Plant PhysiologyDocument170 pagesHawassa University Daye Campus Plant PhysiologySekste MaludaNo ratings yet
- Science 8 Fourth 4 DaysDocument4 pagesScience 8 Fourth 4 DaysAshley Lopez100% (5)
- BT1001 Biology For Engineers PDFDocument2 pagesBT1001 Biology For Engineers PDFsanthi saranyaNo ratings yet
- Role of Jalauka in Diabetic Foot UlcerDocument12 pagesRole of Jalauka in Diabetic Foot UlcerMinakshi Gulshan SharmaNo ratings yet
- The Brain Has A Body Adaptive Behavior Emerges From Interactions of Nervous System, Body and Environment by Hillel J. Chiel and Randall D. Beer 1997Document5 pagesThe Brain Has A Body Adaptive Behavior Emerges From Interactions of Nervous System, Body and Environment by Hillel J. Chiel and Randall D. Beer 1997AmiraNo ratings yet
- I. Clinical Summary A. General Data ProfileDocument8 pagesI. Clinical Summary A. General Data ProfileDayan CabrigaNo ratings yet
- In Focus Periodic Paralysis PDFDocument8 pagesIn Focus Periodic Paralysis PDFLisa Pratiwi RNo ratings yet
- NAPLEX Prep 2018 Ch01 CardioDocument56 pagesNAPLEX Prep 2018 Ch01 CardioFaith50% (2)
- rr420403 Bio Medical InstrumentationDocument4 pagesrr420403 Bio Medical InstrumentationSRINIVASA RAO GANTANo ratings yet
- Practice Test 2 1st Sem 2013Document11 pagesPractice Test 2 1st Sem 2013Isabel Barredo Del MundoNo ratings yet
- Chapter 9 - Transport in AnimalsDocument6 pagesChapter 9 - Transport in AnimalsLeann LeeNo ratings yet
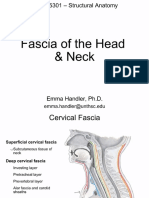
- Fascia of The Head & NeckDocument8 pagesFascia of The Head & NeckMahesh Jung KhadkaNo ratings yet
- Information For You: An Ectopic PregnancyDocument25 pagesInformation For You: An Ectopic PregnancyAnonymous layszXmsNo ratings yet
- Valorization of CoffeeDocument9 pagesValorization of CoffeeEditor IJTSRDNo ratings yet
- Napfa Resource Book For Pe Teachers 20140506Document25 pagesNapfa Resource Book For Pe Teachers 20140506api-259265137100% (1)
- Ventriculo UnicoDocument19 pagesVentriculo UnicoDavid MartínezNo ratings yet
- Interstitial Fluid Flow Induces Myofibroblast Differentiation and Collagen Alignment in VitroDocument9 pagesInterstitial Fluid Flow Induces Myofibroblast Differentiation and Collagen Alignment in VitroPedro FonsecaNo ratings yet
- Anatomy McqsDocument2 pagesAnatomy McqsNoor Saleem100% (2)
- Congenital HypothyroidismDocument21 pagesCongenital HypothyroidismIkhsan Nurkholis SudradjatNo ratings yet
- Urinalysis 2Document9 pagesUrinalysis 2Dawn GonzagaNo ratings yet
- SOAP - Case Study FormatsDocument8 pagesSOAP - Case Study FormatsDr-Sanjay SinghaniaNo ratings yet
- Mcqs On Excretory SystemDocument3 pagesMcqs On Excretory SystemIshfaq LoneNo ratings yet
- Medical Supplies - English Vocabulary: Term MeaningDocument16 pagesMedical Supplies - English Vocabulary: Term MeaningedonaNo ratings yet
- Neuromuscular Taping For The Upper LimbDocument4 pagesNeuromuscular Taping For The Upper LimbFayza RihastaraNo ratings yet
- Umm Al-Qura University Faculty of PharmacyDocument168 pagesUmm Al-Qura University Faculty of Pharmacyapi-19793040No ratings yet
- Fire in The Water Rising Water Vampire Diaries FanfictionDocument65 pagesFire in The Water Rising Water Vampire Diaries FanfictionCristina Luntraru0% (1)
- Neurological Complications in Lumbar Spine SurgeryDocument9 pagesNeurological Complications in Lumbar Spine SurgeryMimi SyakilaNo ratings yet
- Jurnal 11Document14 pagesJurnal 11Zulvi RiriNo ratings yet
- Principles of Vegetative PropagationDocument5 pagesPrinciples of Vegetative PropagationJensenNo ratings yet
- Atika School-5172016 - Biology Form 2Document14 pagesAtika School-5172016 - Biology Form 2DenisNo ratings yet