You might also like
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (587)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (890)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (265)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- CretDocument63 pagesCretMtpa MashoorNo ratings yet
- The Function of Vitamins and Mineral in Hair LossDocument2 pagesThe Function of Vitamins and Mineral in Hair LossDian AcilNo ratings yet
- Detailed Lesson Plan in Quarter 4 WK 1 Digestive ProcessDocument10 pagesDetailed Lesson Plan in Quarter 4 WK 1 Digestive ProcessmalouNo ratings yet
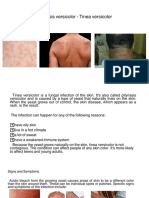
- Pityriasis VersicolorDocument6 pagesPityriasis Versicolorh8j5fnyh7dNo ratings yet
- Ebr MacalolotDocument4 pagesEbr Macalolotquen.chuaNo ratings yet
- ADR UzbekistanDocument80 pagesADR UzbekistanJose Juan Velazquez GarciaNo ratings yet
- JKNKLKLDocument10 pagesJKNKLKLCyntia AndrinaNo ratings yet
- Homeoapthy For InfantsDocument9 pagesHomeoapthy For Infantspavani_upNo ratings yet
- Bhs Inggris - Health and Nursing Related Vocabulary-1Document9 pagesBhs Inggris - Health and Nursing Related Vocabulary-1septynovitaNo ratings yet
- The Herbal of Dioscorides Materia MedicaDocument78 pagesThe Herbal of Dioscorides Materia MedicaShari Cook100% (2)
- Double Unders MasteryDocument13 pagesDouble Unders MasterySebastian Hurtado100% (2)
- Abdominal ExaminationDocument14 pagesAbdominal ExaminationValeria Guerra CastilloNo ratings yet
- Material Safety Data Sheet: 1. Chemical Product and Company IdentificationDocument7 pagesMaterial Safety Data Sheet: 1. Chemical Product and Company IdentificationKun Adi ReksatamaNo ratings yet
- DLL - Fourth GradingDocument5 pagesDLL - Fourth GradingMeek Adan RodriguezNo ratings yet
- Anticoagulation Options For Intermittent HDDocument10 pagesAnticoagulation Options For Intermittent HDNarinder SharmaNo ratings yet
- ShaylaDocument3 pagesShaylaapi-530728661No ratings yet
- Headache - 2001 - Jacome - Transitional Interpersonality Thunderclap HeadacheDocument4 pagesHeadache - 2001 - Jacome - Transitional Interpersonality Thunderclap HeadacheactualashNo ratings yet
- Understanding Cholesterol Levels and Risks: A Guide to Lipid ProfilesDocument48 pagesUnderstanding Cholesterol Levels and Risks: A Guide to Lipid ProfilesAnonymous o3Q3QXENo ratings yet
- Medications and Antidotes ChartDocument2 pagesMedications and Antidotes ChartkNo ratings yet
- Krok 2 - 2021 - 26 JulyDocument19 pagesKrok 2 - 2021 - 26 JulyNicole VinnikNo ratings yet
- Gtbr2016 Main TextDocument214 pagesGtbr2016 Main TextRaimundo Isidro MachavaNo ratings yet
- How Can Women Make The RulesDocument0 pagesHow Can Women Make The Ruleshme_s100% (2)
- Biochem 2 Chapter 1Document19 pagesBiochem 2 Chapter 1Suzaila's SyfaNo ratings yet
- Application of Roy TheoryDocument32 pagesApplication of Roy TheoryAnusha VergheseNo ratings yet
- Stallard's eye surgery pdf free download guideDocument3 pagesStallard's eye surgery pdf free download guideCod Mobile100% (1)
- Chapter II-Vocabulary ExerciseDocument3 pagesChapter II-Vocabulary ExerciseJessica KarinaNo ratings yet
- Understand Glaucoma: Causes, Diagnosis and TreatmentDocument57 pagesUnderstand Glaucoma: Causes, Diagnosis and TreatmentDarshan SinghNo ratings yet
- Tumor Detection Through Mri Brain Images: Rohit Arya 20MCS1009Document25 pagesTumor Detection Through Mri Brain Images: Rohit Arya 20MCS1009Rohit AryaNo ratings yet
- A Novel Lymphocyte Transformation Test (LTT-MELISAR) For Lyme BorreliosisDocument8 pagesA Novel Lymphocyte Transformation Test (LTT-MELISAR) For Lyme Borreliosisdebnathsuman49No ratings yet
- Atopic Eczema in Adulthood and Risk of Depression and Anxiety: A Population-Based Cohort StudyDocument26 pagesAtopic Eczema in Adulthood and Risk of Depression and Anxiety: A Population-Based Cohort Studyyenny handayani sihiteNo ratings yet