You might also like
- Ulok Dr. Rina 18.01 22.00Document84 pagesUlok Dr. Rina 18.01 22.00Cindy PrayogoNo ratings yet
- ENT Dr. Angel Case 29 Jan 17.00Document85 pagesENT Dr. Angel Case 29 Jan 17.00Cindy PrayogoNo ratings yet
- Case Analysis DR RinaDocument30 pagesCase Analysis DR RinaCindy PrayogoNo ratings yet
- Case Report Intermittent Asthma, Allergic Rhinitis, Obesity in A ChildDocument91 pagesCase Report Intermittent Asthma, Allergic Rhinitis, Obesity in A ChildCindy PrayogoNo ratings yet
- Case Report Allergic RhinitisDocument67 pagesCase Report Allergic RhinitisCindy PrayogoNo ratings yet
- Ovoran Nicu 2 Kamis, 1 April 2021: GDS/24 JamDocument1 pageOvoran Nicu 2 Kamis, 1 April 2021: GDS/24 JamCindy PrayogoNo ratings yet
- 1 April 2021Document1 page1 April 2021Cindy PrayogoNo ratings yet
- Ec ABO Inkomptabilitas: Christine, S.Ked 171 2005 0035Document1 pageEc ABO Inkomptabilitas: Christine, S.Ked 171 2005 0035Cindy PrayogoNo ratings yet
- CR HSPDocument55 pagesCR HSPCindy PrayogoNo ratings yet
- Instructors: First Aid TeamDocument6 pagesInstructors: First Aid TeamCindy PrayogoNo ratings yet
- CSF and Cerebral Blood FlowDocument58 pagesCSF and Cerebral Blood FlowCindy PrayogoNo ratings yet
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (400)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (345)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (121)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- Laban - Sub Kamal Salman Mahmoud Jaber Mustafa: Done byDocument40 pagesLaban - Sub Kamal Salman Mahmoud Jaber Mustafa: Done byKool Kaish100% (1)
- Essential Infantile Esotropia An Unusual Case ReportDocument3 pagesEssential Infantile Esotropia An Unusual Case ReportInternational Journal of Innovative Science and Research TechnologyNo ratings yet
- La Multisensorialidad Del Sabor PDFDocument4 pagesLa Multisensorialidad Del Sabor PDF690218No ratings yet
- Sumi - Presentation On BlindnessDocument23 pagesSumi - Presentation On Blindnessabhigyan priyadarshanNo ratings yet
- Sala Hall/Recepción Sala A // Auditorio Subsuelo Sala B // SUM 5° PISO Sala C // Entrada Auditorio Inscripción Información (Hall/Recepción)Document2 pagesSala Hall/Recepción Sala A // Auditorio Subsuelo Sala B // SUM 5° PISO Sala C // Entrada Auditorio Inscripción Información (Hall/Recepción)Lucho BenottoNo ratings yet
- Hearing ImpaimentDocument29 pagesHearing ImpaimentAPRILYN REFENDOR BCAEd - 4DNo ratings yet
- Clinical Exam For OphthalmologyDocument6 pagesClinical Exam For Ophthalmologynuuraaisya50% (2)
- COPIC Color ChartDocument1 pageCOPIC Color ChartEstephani Cal DerNo ratings yet
- Mathematics in Medicine and Medical ScienceDocument10 pagesMathematics in Medicine and Medical ScienceFF EDITZNo ratings yet
- Theories of Colour VisionDocument4 pagesTheories of Colour VisionZumairi Tamin0% (1)
- CATARACTDocument61 pagesCATARACTMukta KadamNo ratings yet
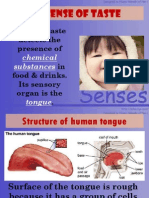
- 1.4 Sense of Taste: Sense of Taste Detects The Presence of in Food & Drinks. Its Sensory Organ Is TheDocument6 pages1.4 Sense of Taste: Sense of Taste Detects The Presence of in Food & Drinks. Its Sensory Organ Is TheMissz WazeeNo ratings yet
- The Basics of Retinal OCT Oct-BDocument58 pagesThe Basics of Retinal OCT Oct-BJomarNo ratings yet
- Anatomyofthevestibularsystem Areview PDFDocument8 pagesAnatomyofthevestibularsystem Areview PDFJacqueline Rebolledo BravoNo ratings yet
- The Senses. Comprehensive Reference (NeuroscienDocument4,112 pagesThe Senses. Comprehensive Reference (NeuroscienIndira EspitiaNo ratings yet
- Color Theory Liquisoft ComDocument1 pageColor Theory Liquisoft Comsilent_pkNo ratings yet
- Retinal Detachment UncenDocument38 pagesRetinal Detachment UncenNixal KurniawanNo ratings yet
- UGH Syndrome After Lens Implantation PDFDocument2 pagesUGH Syndrome After Lens Implantation PDFkameliasitorusNo ratings yet
- Color Theory and Its Application in Art and Design PDFDocument303 pagesColor Theory and Its Application in Art and Design PDFSaksham Kulpati100% (3)
- Accommodative EsotropiaDocument3 pagesAccommodative EsotropiaMedi Sinabutar100% (1)
- ImageryDocument35 pagesImageryJose Ramon CarlosNo ratings yet
- LG 55LM6700 CNET Review Calibration ResultsDocument7 pagesLG 55LM6700 CNET Review Calibration ResultsDavid KatzmaierNo ratings yet
- Anatomy of Human EarDocument5 pagesAnatomy of Human EarSharanz Twentyfour SevenNo ratings yet
- Refractive ErrorsDocument7 pagesRefractive ErrorsJim Jose AntonyNo ratings yet
- 21st Century Module 8 - PDFDocument12 pages21st Century Module 8 - PDFJoey Hermusora Espanto100% (12)
- Visual ImpairmentDocument47 pagesVisual Impairmentcj romagosa67% (3)
- Preparation: Competency Performance Checklist Assessing Ears Student: Date: - Skill Yes NoDocument2 pagesPreparation: Competency Performance Checklist Assessing Ears Student: Date: - Skill Yes NoCyril Joy N. FernandoNo ratings yet
- Case Study GeriatricDocument4 pagesCase Study GeriatricJoshua HavanaNo ratings yet
- Vision Training Program - DR - GalaDocument150 pagesVision Training Program - DR - Galaarvindhans100% (2)
- Rajiv Dhawan ENT Ear Test Post ClassDocument92 pagesRajiv Dhawan ENT Ear Test Post Classdk46dg5j6vNo ratings yet