You might also like
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- Natural Treatment of Lung Disease, COPD, Asthma, Bronchitis, SCIO Eductor StudyDocument121 pagesNatural Treatment of Lung Disease, COPD, Asthma, Bronchitis, SCIO Eductor StudyratudobitsururNo ratings yet
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (587)
- Project Report On Indian BirdsDocument9 pagesProject Report On Indian BirdsMridul kejriwal50% (2)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (894)
- COPD - Case PresentationDocument68 pagesCOPD - Case PresentationJaslir MendozaNo ratings yet
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
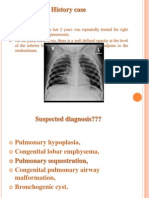
- Congenital Lung MalformationsDocument115 pagesCongenital Lung MalformationsAhmad Abu KushNo ratings yet
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- Pleural EffusionDocument24 pagesPleural Effusionwheeyycoldandhot55No ratings yet
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- Krok 2 2011 + AnswersDocument69 pagesKrok 2 2011 + AnswersKrish RoopunNo ratings yet
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- Measure Your Lung Capacity with Fun Science ActivitiesDocument15 pagesMeasure Your Lung Capacity with Fun Science ActivitiesSthephen MarquezNo ratings yet
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Pathology of The Lungs 3rd Ed (PDF) (Tahir99) VRGDocument746 pagesPathology of The Lungs 3rd Ed (PDF) (Tahir99) VRGCosmin SilviuNo ratings yet
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Respiratory SystemDocument13 pagesRespiratory SystemUsman WaheedNo ratings yet
- Block 2Document14 pagesBlock 2PaolaNo ratings yet
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Serrapeptase - Benefits, Dosage, Dangers, and Side EffectsDocument11 pagesSerrapeptase - Benefits, Dosage, Dangers, and Side EffectsHansley Templeton Cook100% (1)
- Sage GuideDocument236 pagesSage GuideArryanNo ratings yet
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- Diagnosis Banding Ground Glass Opacities Pada Ct-Scan Toraks Di Era Covid 19Document22 pagesDiagnosis Banding Ground Glass Opacities Pada Ct-Scan Toraks Di Era Covid 19rbatjun576No ratings yet
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- Burned Bodies Post-Mortem Computed Tomography PDFDocument13 pagesBurned Bodies Post-Mortem Computed Tomography PDFNabila Noor IswariNo ratings yet
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- WS Chapter 9Document24 pagesWS Chapter 9Jason Timothy KoesmanggalaNo ratings yet
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Gas Exchange in Humans MYP4 BiologyDocument52 pagesGas Exchange in Humans MYP4 Biologydiyar alassafNo ratings yet
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Risks of SmokingDocument10 pagesRisks of SmokingJeravem OrtiyasNo ratings yet
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (265)
- Fresh Fruit and Vegetable Juices by WalkerDocument96 pagesFresh Fruit and Vegetable Juices by WalkerJo Doe100% (4)
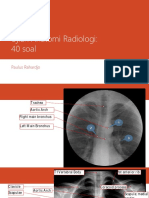
- Ujian Anatomi Radiologi: 40 Soal: Paulus RahardjoDocument16 pagesUjian Anatomi Radiologi: 40 Soal: Paulus RahardjoEga Candra FaurizaNo ratings yet
- Obstructive and restrictive lung diseases and lung tumorsDocument6 pagesObstructive and restrictive lung diseases and lung tumorsZagros MotamedNo ratings yet
- The Human Body in Health and Illness e Book 6th Edition Ebook PDFDocument61 pagesThe Human Body in Health and Illness e Book 6th Edition Ebook PDFcraig.suarez45797% (34)
- PCAP-c Group A NiggasDocument39 pagesPCAP-c Group A NiggasJenny YenNo ratings yet
- MSDSJJDocument1,078 pagesMSDSJJrossangel35No ratings yet
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Free Report - Orofacial Exercises To Cure Sleep ApneaDocument16 pagesFree Report - Orofacial Exercises To Cure Sleep Apneaivan2504100% (2)
- Spirometry: Predicting Risk and Outcome: Alessandro Brunelli, MD, Gaetano Rocco, MD, FRCSDocument8 pagesSpirometry: Predicting Risk and Outcome: Alessandro Brunelli, MD, Gaetano Rocco, MD, FRCSAfien MuktiNo ratings yet
- ANATOMY AND PHYSIOLOGY-flashcardsDocument21 pagesANATOMY AND PHYSIOLOGY-flashcardsJohn Wayne PharrNo ratings yet
- Pulmonary Rehabilitation Slide PresentationDocument24 pagesPulmonary Rehabilitation Slide Presentationdranoj0070% (1)
- PMRDocument17 pagesPMRJoy SinghNo ratings yet
- CT ChestDocument277 pagesCT ChestpraveenbhavniNo ratings yet
- Direction: Encircle The Letter of The Correct AnswerDocument2 pagesDirection: Encircle The Letter of The Correct Answerariel agos100% (1)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)