You might also like
- Camp's Zoology by the Numbers: A comprehensive study guide in outline form for advanced biology courses, including AP, IB, DE, and college courses.From EverandCamp's Zoology by the Numbers: A comprehensive study guide in outline form for advanced biology courses, including AP, IB, DE, and college courses.No ratings yet
- Pattern Formation and Growth During Floral OrganogDocument9 pagesPattern Formation and Growth During Floral OrganogSweeti MandalNo ratings yet
- Baroux 2004 Pla Nuclear Fusions Contrib To Polyploid of Gigantic Nuclei in Chalaz Endosp of atDocument9 pagesBaroux 2004 Pla Nuclear Fusions Contrib To Polyploid of Gigantic Nuclei in Chalaz Endosp of atAJMRNo ratings yet
- The Transient Cortical Zone in The Adrenal Gland: The Mystery of The Adrenal X-ZoneDocument13 pagesThe Transient Cortical Zone in The Adrenal Gland: The Mystery of The Adrenal X-ZoneCandle Jo'inNo ratings yet
- Germline Regulatory Element of Oct-4 Specific For The Totipotent Cycle ofDocument14 pagesGermline Regulatory Element of Oct-4 Specific For The Totipotent Cycle ofMariane Gabriela Cesar Ribeiro FerreiraNo ratings yet
- Biología Del Cuerpo LuteoDocument12 pagesBiología Del Cuerpo LuteoVioleta v cNo ratings yet
- Genetics of Axis Specification in DrosophilaDocument3 pagesGenetics of Axis Specification in DrosophilaJohnkevin NocheNo ratings yet
- D MelanogasterDocument40 pagesD Melanogastersunifeb128075No ratings yet
- Paternal Investment and Lntracellular Sperm-Egg Interactions During and Following Fer T Zat On inDocument27 pagesPaternal Investment and Lntracellular Sperm-Egg Interactions During and Following Fer T Zat On inじょしら フィアンナNo ratings yet
- As Epidermal Stem Cells Age They Do Not Substantially Change Their CharacteristicsDocument9 pagesAs Epidermal Stem Cells Age They Do Not Substantially Change Their CharacteristicsMichał PikułaNo ratings yet
- Panfilio, 2006 KatatrepsisDocument18 pagesPanfilio, 2006 KatatrepsisGabriel BuonoNo ratings yet
- Drosophila Embryogenesis: The Formation of Fruit Fly EmbryosDocument6 pagesDrosophila Embryogenesis: The Formation of Fruit Fly EmbryosVikram Pkb SharmaNo ratings yet
- 1519024439M06AxisspecificationindrosophilaQuad1Document18 pages1519024439M06AxisspecificationindrosophilaQuad1Mahendra KatreNo ratings yet
- Southworth Et Al 2001-Male GametogenesisDocument16 pagesSouthworth Et Al 2001-Male GametogenesisAde MulyanaNo ratings yet
- Lavdas Et Al. - J.neurosc. - 1999 - The MGE Gives Rise To A Population of Early Neurons in The Developing Cerebral CortexDocument8 pagesLavdas Et Al. - J.neurosc. - 1999 - The MGE Gives Rise To A Population of Early Neurons in The Developing Cerebral CortexCris AdFNo ratings yet
- A Drosophila Gain-of-Function Screen For Candidate Genes Involved in Steroid-Dependent Neuroendocrine Cell RemodelingDocument19 pagesA Drosophila Gain-of-Function Screen For Candidate Genes Involved in Steroid-Dependent Neuroendocrine Cell RemodelingLautaro BelfioriNo ratings yet
- 1998 Egfr GfapDocument13 pages1998 Egfr GfapManuelNo ratings yet
- tmpF930 TMPDocument21 pagestmpF930 TMPFrontiersNo ratings yet
- Hal 277 - 309Document41 pagesHal 277 - 309Baiq Indah Komala SariNo ratings yet
- Looking at The Big Picture Understanding How The Oviduct's Dialogue With Gametes and TheDocument14 pagesLooking at The Big Picture Understanding How The Oviduct's Dialogue With Gametes and TheFabiane MoraesNo ratings yet
- Bangs 2000Document12 pagesBangs 2000laciyeg352No ratings yet
- Baroux 2007 PLC The 3n Endosperm Genome of at Adopts A Peculiar Parental-dos-Dep Chrom OrganizDocument13 pagesBaroux 2007 PLC The 3n Endosperm Genome of at Adopts A Peculiar Parental-dos-Dep Chrom OrganizAJMRNo ratings yet
- Example Lab ReportDocument20 pagesExample Lab ReportbobtentpegNo ratings yet
- The Avian OrganizerDocument7 pagesThe Avian OrganizerTamara OliveiraNo ratings yet
- AK MergedDocument37 pagesAK MergedAaryan GuptaNo ratings yet
- Medical Journal of Cell Biology Drosophila Melanogaster Research History, Breakthrough and PerspectivesDocument4 pagesMedical Journal of Cell Biology Drosophila Melanogaster Research History, Breakthrough and PerspectivesIgor DemićNo ratings yet
- Cloning Human BeingsDocument312 pagesCloning Human BeingswalterkondorNo ratings yet
- #Domínguez 1997Document12 pages#Domínguez 1997yustica arisnaNo ratings yet
- Caenorhabditis gonadeDocument21 pagesCaenorhabditis gonadeIstván NagyNo ratings yet
- Shiang 1994Document8 pagesShiang 1994Daniela NarvaezNo ratings yet
- Stem Cell FactorDocument22 pagesStem Cell FactoractivnetNo ratings yet
- Gastrulation in the Mouse EmbryoDocument28 pagesGastrulation in the Mouse EmbryoDIEGO FERNANDO URIBE JARAMILLONo ratings yet
- Paper Neupogen PDFDocument13 pagesPaper Neupogen PDFFranNo ratings yet
- Journal of Anatomy - 2003 - Hill - Sonic hedgehog restricted expression and limb dysmorphologiesDocument8 pagesJournal of Anatomy - 2003 - Hill - Sonic hedgehog restricted expression and limb dysmorphologiesJPNo ratings yet
- (Vincent Et Al, 1997) Ntegration of The Head and Trunk Segmentation Systems Controls Cephalic Furrow Formation in DrosophilaDocument8 pages(Vincent Et Al, 1997) Ntegration of The Head and Trunk Segmentation Systems Controls Cephalic Furrow Formation in DrosophilaエリタノアンソネーNo ratings yet
- Roote Prokop SupplMat 1v3.3Document35 pagesRoote Prokop SupplMat 1v3.3Diana BicazanNo ratings yet
- The Onset of Germ Cell Migration in The Mouse EmbryoDocument8 pagesThe Onset of Germ Cell Migration in The Mouse EmbryoLin Zahir González GonzálezNo ratings yet
- Molecular Genetic Studies Confirm The Role of Brassinosteroids in Plant Growth and DevelopmentDocument8 pagesMolecular Genetic Studies Confirm The Role of Brassinosteroids in Plant Growth and DevelopmentBintang AlfaenNo ratings yet
- Cfos, Jun y ExploracionDocument12 pagesCfos, Jun y ExploracionLuisa Fernanda Acevedo ReyesNo ratings yet
- Cell Cycle in Mouse Development: Maria A Ciemerych and Peter SicinskiDocument22 pagesCell Cycle in Mouse Development: Maria A Ciemerych and Peter SicinskiJamilla Ann Fatima FelicianoNo ratings yet
- Molecular Embryology of The ForegutDocument4 pagesMolecular Embryology of The Foregutproject-247758No ratings yet
- Molecular Embryology of The ForegutDocument4 pagesMolecular Embryology of The ForegutmikeNo ratings yet
- Molecular Embryology of The ForegutDocument4 pagesMolecular Embryology of The ForegutmikeNo ratings yet
- Molecular Embryology of The ForegutDocument4 pagesMolecular Embryology of The ForegutmikeNo ratings yet
- Embryonic Expression of The Divergent Drosophila Isoform Is Required For Larval BehaviorDocument11 pagesEmbryonic Expression of The Divergent Drosophila Isoform Is Required For Larval BehaviorGustavo FelpeNo ratings yet
- 1 s2.0 S2667290121000310 MainDocument11 pages1 s2.0 S2667290121000310 MainIdeologiesNo ratings yet
- Formation Transcription Switches For Protoxylem and Metaxylem VesselDocument7 pagesFormation Transcription Switches For Protoxylem and Metaxylem VesselMutia Diansafitri SuhudNo ratings yet
- Regional Specification in The Early Embryo of The Brittle Star Ophiopholis AculeataDocument16 pagesRegional Specification in The Early Embryo of The Brittle Star Ophiopholis AculeataJoElunico QuezadaNo ratings yet
- Drosophila Yurt Is A New Protein-4.1-Like Protein Required For Epithelial MorphogenesisDocument9 pagesDrosophila Yurt Is A New Protein-4.1-Like Protein Required For Epithelial MorphogenesismilicmilNo ratings yet
- Crozatier Et Al-2007-Cellular MicrobiologyDocument10 pagesCrozatier Et Al-2007-Cellular MicrobiologyVINEET KUREELNo ratings yet
- Chapter 17 - Endocrine Systems - 2020 - The Zebrafish in Biomedical ResearchDocument15 pagesChapter 17 - Endocrine Systems - 2020 - The Zebrafish in Biomedical ResearchNicolas BaronNo ratings yet
- Genetic Control of Development: DifferentiationDocument4 pagesGenetic Control of Development: DifferentiationElizabeth Valle EspinozaNo ratings yet
- Transgenic AnimalsDocument12 pagesTransgenic AnimalsAbhimanyu JainNo ratings yet
- Developmental Regulation of The Hox Genes During Axial Morphogenesis in The MouseDocument16 pagesDevelopmental Regulation of The Hox Genes During Axial Morphogenesis in The MouseVero Claros TapiaNo ratings yet
- Kura Ku 2008Document13 pagesKura Ku 2008J.D. NobleNo ratings yet
- Morphological Transitions and The Genetic Basis of The Evolution of Extraembryonic Tissues in FliesDocument123 pagesMorphological Transitions and The Genetic Basis of The Evolution of Extraembryonic Tissues in FliesMatteen RafiqiNo ratings yet
- TMP 9 E37Document8 pagesTMP 9 E37FrontiersNo ratings yet
- Rougier 2001 Pathenogenisis in MammalsDocument7 pagesRougier 2001 Pathenogenisis in MammalsFroyNo ratings yet
- Chem 9Document7 pagesChem 9Cao Thị Vân GiangNo ratings yet
- 5 Techniques for inducing transgenics in fishDocument1 page5 Techniques for inducing transgenics in fishLaleth Mendoza OjalesNo ratings yet
- Drosophila Yurt Is A New Protein-4.1-Like Protein Required For Epithelial MorphogenesisDocument9 pagesDrosophila Yurt Is A New Protein-4.1-Like Protein Required For Epithelial MorphogenesismilicmilNo ratings yet
- IzadoostDocument5 pagesIzadoostmilicmilNo ratings yet
- Godt TepassDocument5 pagesGodt TepassmilicmilNo ratings yet
- Pax7 - 2Document11 pagesPax7 - 2milicmilNo ratings yet
- IzadoostDocument5 pagesIzadoostmilicmilNo ratings yet
- Myosin II PaperDocument13 pagesMyosin II PapermilicmilNo ratings yet
- Transport Phenomena in Nervous System PDFDocument538 pagesTransport Phenomena in Nervous System PDFUdayanidhi RNo ratings yet
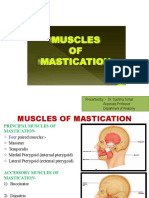
- Presented By:-Dr. Sushma Tomar Associate Professor Department of AnatomyDocument20 pagesPresented By:-Dr. Sushma Tomar Associate Professor Department of AnatomyCristine EchaveNo ratings yet
- Perkecambahan Dan Pertumbuhan Palem Jepang (Actinophloeus Macarthurii Becc.) Akibat Perendaman Biji Dalam LumpurDocument7 pagesPerkecambahan Dan Pertumbuhan Palem Jepang (Actinophloeus Macarthurii Becc.) Akibat Perendaman Biji Dalam LumpurNurma YahyaNo ratings yet
- Agilent ASMS 2019 ThP762 PosterDocument4 pagesAgilent ASMS 2019 ThP762 PostersppNo ratings yet
- Amazing Facts - Exercises 1Document3 pagesAmazing Facts - Exercises 1Alina DorobatNo ratings yet
- Ewh Ix PDFDocument80 pagesEwh Ix PDFOR Premium FreeNo ratings yet
- Chapter 2 Medical Terminology Verified AnswersDocument5 pagesChapter 2 Medical Terminology Verified AnswersGregg ProducerNo ratings yet
- Poultry Science MCQDocument12 pagesPoultry Science MCQelanthamizhmaran83% (6)
- Hemostatic Effects of Peperomia PellucidDocument41 pagesHemostatic Effects of Peperomia PellucidGina Rose De GuzmanNo ratings yet
- CHANGES IN ANTI OXIDANT ENZYME ACTIVITIES IN Pseudomonas Syringae PV Syringae (BACTERIAL BROWN SPOT) INFECTED SEEDLINGS OF Vigna Radiata L.Document5 pagesCHANGES IN ANTI OXIDANT ENZYME ACTIVITIES IN Pseudomonas Syringae PV Syringae (BACTERIAL BROWN SPOT) INFECTED SEEDLINGS OF Vigna Radiata L.ijsretNo ratings yet
- Diencephalon PresentationDocument19 pagesDiencephalon Presentation300rNo ratings yet
- Psychiatric Mental Health Nursing Concepts of Care in Evidence Based Practice 8th Edition Townsend Test BankDocument36 pagesPsychiatric Mental Health Nursing Concepts of Care in Evidence Based Practice 8th Edition Townsend Test Bankquemefuloathableilljzf100% (26)
- Sample Lab ReportDocument7 pagesSample Lab ReportPutri Syalieyana0% (1)
- The Story of OrchidDocument7 pagesThe Story of OrchidLilian Kimwell SastadoNo ratings yet
- Summary of Qualitative Tests (Pharmacognosy)Document8 pagesSummary of Qualitative Tests (Pharmacognosy)kidsaintfineNo ratings yet
- TC QMM 56942Document120 pagesTC QMM 56942Fernando R EpilNo ratings yet
- Effects of Sleep Deprivation among Students in Saint Francis of Assisi CollegeDocument50 pagesEffects of Sleep Deprivation among Students in Saint Francis of Assisi College• Cielo •No ratings yet
- This Topic. So Congratulations Because Today I'm Going To Talk About TheDocument3 pagesThis Topic. So Congratulations Because Today I'm Going To Talk About TheJoon Bok NamleeNo ratings yet
- Flowering PlantsDocument43 pagesFlowering Plantskingbanakon100% (1)
- Ssac 2104Document100 pagesSsac 2104Amrutha amrNo ratings yet
- Evolution of Pharmacy in the PhilippinesDocument2 pagesEvolution of Pharmacy in the PhilippinesRosemarie OngNo ratings yet
- MDU Open Elective 3rd Sem Date Sheet May 2019Document1 pageMDU Open Elective 3rd Sem Date Sheet May 2019Ratan DeshNo ratings yet
- Protochordata FIXDocument33 pagesProtochordata FIXSylvia AnggraeniNo ratings yet
- Egg Quality Manual Web PDFDocument44 pagesEgg Quality Manual Web PDFVeterinario La EsperanzaNo ratings yet
- Cell Membrane: A Selective BarrierDocument23 pagesCell Membrane: A Selective BarrierHama JamalNo ratings yet
- 421 Reproduction Developmental Toxicity Screening TestDocument10 pages421 Reproduction Developmental Toxicity Screening TestAshishNo ratings yet
- Turbidimetry and Nephelometry1Document3 pagesTurbidimetry and Nephelometry1maxim_crank6101100% (1)
- MOKUSAKU APPLICATIONS (Version 1)Document1 pageMOKUSAKU APPLICATIONS (Version 1)api-19621568No ratings yet
- Conference on Global Issues in Agricultural, Environmental and Forestry SciencesDocument16 pagesConference on Global Issues in Agricultural, Environmental and Forestry SciencesShirish NarnawareNo ratings yet
- Ap Environmental Science Free-Response QuestionsDocument6 pagesAp Environmental Science Free-Response Questionsapi-234419767No ratings yet
- Why We Die: The New Science of Aging and the Quest for ImmortalityFrom EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityRating: 3.5 out of 5 stars3.5/5 (2)
- 10% Human: How Your Body's Microbes Hold the Key to Health and HappinessFrom Everand10% Human: How Your Body's Microbes Hold the Key to Health and HappinessRating: 4 out of 5 stars4/5 (33)
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceFrom EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceRating: 4.5 out of 5 stars4.5/5 (515)
- The Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindFrom EverandThe Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindRating: 4.5 out of 5 stars4.5/5 (93)
- Crypt: Life, Death and Disease in the Middle Ages and BeyondFrom EverandCrypt: Life, Death and Disease in the Middle Ages and BeyondRating: 4 out of 5 stars4/5 (3)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisFrom EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisRating: 3.5 out of 5 stars3.5/5 (2)
- This Is Your Brain On Parasites: How Tiny Creatures Manipulate Our Behavior and Shape SocietyFrom EverandThis Is Your Brain On Parasites: How Tiny Creatures Manipulate Our Behavior and Shape SocietyRating: 3.5 out of 5 stars3.5/5 (31)
- The Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionFrom EverandThe Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionRating: 4 out of 5 stars4/5 (811)
- All That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesFrom EverandAll That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesRating: 4.5 out of 5 stars4.5/5 (396)
- The Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorFrom EverandThe Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorNo ratings yet
- Wayfinding: The Science and Mystery of How Humans Navigate the WorldFrom EverandWayfinding: The Science and Mystery of How Humans Navigate the WorldRating: 4.5 out of 5 stars4.5/5 (18)
- Undeniable: How Biology Confirms Our Intuition That Life Is DesignedFrom EverandUndeniable: How Biology Confirms Our Intuition That Life Is DesignedRating: 4 out of 5 stars4/5 (11)
- The Lives of Bees: The Untold Story of the Honey Bee in the WildFrom EverandThe Lives of Bees: The Untold Story of the Honey Bee in the WildRating: 4.5 out of 5 stars4.5/5 (44)
- The Second Brain: A Groundbreaking New Understanding of Nervous Disorders of the Stomach and IntestineFrom EverandThe Second Brain: A Groundbreaking New Understanding of Nervous Disorders of the Stomach and IntestineRating: 4 out of 5 stars4/5 (17)
- Eels: An Exploration, from New Zealand to the Sargasso, of the World's Most Mysterious FishFrom EverandEels: An Exploration, from New Zealand to the Sargasso, of the World's Most Mysterious FishRating: 4 out of 5 stars4/5 (30)
- A Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsFrom EverandA Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsRating: 4.5 out of 5 stars4.5/5 (4)
- Human Errors: A Panorama of Our Glitches, from Pointless Bones to Broken GenesFrom EverandHuman Errors: A Panorama of Our Glitches, from Pointless Bones to Broken GenesRating: 3.5 out of 5 stars3.5/5 (56)
- The Mind & The Brain: Neuroplasticity and the Power of Mental ForceFrom EverandThe Mind & The Brain: Neuroplasticity and the Power of Mental ForceNo ratings yet
- Gathering Moss: A Natural and Cultural History of MossesFrom EverandGathering Moss: A Natural and Cultural History of MossesRating: 4.5 out of 5 stars4.5/5 (347)
- The Dog Who Couldn't Stop Loving: How Dogs Have Captured Our Hearts for Thousands of YearsFrom EverandThe Dog Who Couldn't Stop Loving: How Dogs Have Captured Our Hearts for Thousands of YearsNo ratings yet
- Younger for Life: Feel Great and Look Your Best with the New Science of AutojuvenationFrom EverandYounger for Life: Feel Great and Look Your Best with the New Science of AutojuvenationRating: 4 out of 5 stars4/5 (1)
- Darwin's Dangerous Idea: Evolution and the Meaning of LifeFrom EverandDarwin's Dangerous Idea: Evolution and the Meaning of LifeRating: 4 out of 5 stars4/5 (523)
- Superlative: The Biology of ExtremesFrom EverandSuperlative: The Biology of ExtremesRating: 4.5 out of 5 stars4.5/5 (51)
- Why We Sleep: Unlocking the Power of Sleep and DreamsFrom EverandWhy We Sleep: Unlocking the Power of Sleep and DreamsRating: 4.5 out of 5 stars4.5/5 (2083)
- Lymph & Longevity: The Untapped Secret to HealthFrom EverandLymph & Longevity: The Untapped Secret to HealthRating: 4.5 out of 5 stars4.5/5 (13)