You might also like
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (121)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (400)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5795)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1091)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (345)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Naheed Ali, Moshe Lewis-Understanding Pain - An Introduction For Patients and Caregivers-Rowman & Littlefield Publishers (2015)Document415 pagesNaheed Ali, Moshe Lewis-Understanding Pain - An Introduction For Patients and Caregivers-Rowman & Littlefield Publishers (2015)MaryNo ratings yet
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Floppy BabyDocument13 pagesFloppy BabyJorge JhgNo ratings yet
- Fcps IMMDocument22 pagesFcps IMMHaseeb Rasool0% (1)
- Occams Protocol Cheat SheetDocument10 pagesOccams Protocol Cheat Sheetfnandow4lNo ratings yet
- TuberculosisDocument952 pagesTuberculosiseka kurniawanNo ratings yet
- 14 Health Benefits of Dark ChocolateDocument14 pages14 Health Benefits of Dark ChocolateThavam RatnaNo ratings yet
- Introduction - Spectro-Chrome MetryDocument4 pagesIntroduction - Spectro-Chrome MetryitounosNo ratings yet
- WarmTouch User Manual Wt5200 UsDocument34 pagesWarmTouch User Manual Wt5200 Usnobel0001No ratings yet
- Disruption of Radiologist WorkflowDocument6 pagesDisruption of Radiologist WorkflowChrisNo ratings yet
- Jpts 28 3227 PDFDocument5 pagesJpts 28 3227 PDFphildawsonphotoNo ratings yet
- Pi Is 0025619616308187Document13 pagesPi Is 0025619616308187Anthony M CaballeroNo ratings yet
- Case Presentation On LrtiDocument17 pagesCase Presentation On LrtiNewtan DebNo ratings yet
- Dental Academy 2016Document11 pagesDental Academy 2016Priya SargunanNo ratings yet
- Dyshidrotic Area and Severity IndexDocument5 pagesDyshidrotic Area and Severity IndexJaheh JapihehNo ratings yet
- AsafoetidaDocument2 pagesAsafoetidameet143bmNo ratings yet
- People v. QuianzonDocument3 pagesPeople v. QuianzonACNo ratings yet
- Health 3rd Quarter LessonDocument26 pagesHealth 3rd Quarter LessonJason LaroaNo ratings yet
- Improving The Quality of Health Journalism: When Reliability Meets Engagement, Heini MaksimainenDocument60 pagesImproving The Quality of Health Journalism: When Reliability Meets Engagement, Heini MaksimainenRed Ética Segura - FNPINo ratings yet
- VisionDocument8 pagesVisionDr. Sukhwant SinghNo ratings yet
- 2nd DoseDocument1 page2nd DoseAnkem AdityaNo ratings yet
- DHF 2003Document211 pagesDHF 2003Devvi Riasari BunardiNo ratings yet
- Msds F34avturfsiiDocument10 pagesMsds F34avturfsiiAyat TullahNo ratings yet
- Ambulatory Obstetrics GynecologyDocument4 pagesAmbulatory Obstetrics GynecologyNelson R. PouNo ratings yet
- UK Guidelines On The Management of Variceal Hemorrhage in Cirrhotic PatientsDocument26 pagesUK Guidelines On The Management of Variceal Hemorrhage in Cirrhotic Patientsandus007No ratings yet
- Speaking Short QuestionDocument8 pagesSpeaking Short QuestionpraveshkafleNo ratings yet
- Durian Jelly: An Undergraduate Practical ResearchDocument6 pagesDurian Jelly: An Undergraduate Practical ResearchDOLORFEY L. SUMILE100% (1)
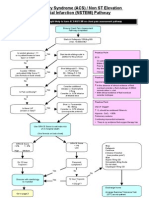
- ACS NSTEMI Clinical PathwayDocument3 pagesACS NSTEMI Clinical PathwayXtiaRNo ratings yet
- Saeed Book Bank: Leading Importers, Exporter, Distributors, Booksellers & Publishers of PakistanDocument15 pagesSaeed Book Bank: Leading Importers, Exporter, Distributors, Booksellers & Publishers of Pakistantalisella50% (2)
- The Effect of The Methanol Extract of Haloxylon Salicornicum (Moq.) Bunge Ex Bioss. On Alleviating in AmmationsDocument8 pagesThe Effect of The Methanol Extract of Haloxylon Salicornicum (Moq.) Bunge Ex Bioss. On Alleviating in AmmationszzakieNo ratings yet
- Combination of Microneedling and 10% Trichloroacetic Acid Peels in The Management of Infraorbital Dark CirclesDocument16 pagesCombination of Microneedling and 10% Trichloroacetic Acid Peels in The Management of Infraorbital Dark CirclesAna Claudia Kordelos DinizNo ratings yet