You might also like
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (265)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (587)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Atlas of Natural CuresDocument515 pagesThe Atlas of Natural CuresChristopher Phillips100% (10)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (894)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- Hormones & Menstruation CycleDocument11 pagesHormones & Menstruation Cyclehiral mistryNo ratings yet
- Male and Female Sex HormonesDocument20 pagesMale and Female Sex HormonesJose Fernando Gregana ElpaNo ratings yet
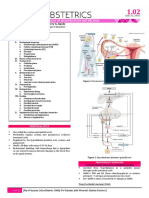
- OB 1 - 1.02 Physiology of Menstruation and Decidua PDFDocument11 pagesOB 1 - 1.02 Physiology of Menstruation and Decidua PDFMDreamerNo ratings yet
- Soal Ujian Stase Fer 5 April 2021Document8 pagesSoal Ujian Stase Fer 5 April 2021Bella DirkNo ratings yet
- Naskah Soal UN Kimia SMA 2014 Paket 1Document15 pagesNaskah Soal UN Kimia SMA 2014 Paket 1Katiman, S.PdNo ratings yet
- Collaborative Learning Enhances Critical ThinkingDocument2 pagesCollaborative Learning Enhances Critical ThinkingMuryati ArchuletaNo ratings yet
- Naskah Soal UN Kimia SMA 2014 Paket 1Document15 pagesNaskah Soal UN Kimia SMA 2014 Paket 1Katiman, S.PdNo ratings yet
- 10 1006@jcis 1996 4564 PDFDocument7 pages10 1006@jcis 1996 4564 PDFMuryati ArchuletaNo ratings yet
- 10 1006@jcis 1996 4564 PDFDocument7 pages10 1006@jcis 1996 4564 PDFMuryati ArchuletaNo ratings yet
- AnastrozoleDocument13 pagesAnastrozoleddandan_2No ratings yet
- Toxicology Report Ethinyl Estradiol: Abbi, Anh, Melody and PaulDocument26 pagesToxicology Report Ethinyl Estradiol: Abbi, Anh, Melody and Paulcachualovely20No ratings yet
- Bo 1994 TheriogenologyDocument16 pagesBo 1994 TheriogenologyGilson Antonio PessoaNo ratings yet
- MethimazoleDocument49 pagesMethimazoleChemistixNo ratings yet
- The Essential Phases of the Menstrual CycleDocument17 pagesThe Essential Phases of the Menstrual CyclePauline Yves IyaNo ratings yet
- The Current Strategy in Hormonal and Non-HormonalDocument25 pagesThe Current Strategy in Hormonal and Non-HormonalJulio YulitaNo ratings yet
- Journal Medicine: The New EnglandDocument7 pagesJournal Medicine: The New EnglandTony RamirezNo ratings yet
- Neural and Hormonal Control of Sexual BehaviorDocument30 pagesNeural and Hormonal Control of Sexual BehaviorKhushi ShuklaNo ratings yet
- Effect of Estrogen and Progesterone On Seed GerminationDocument2 pagesEffect of Estrogen and Progesterone On Seed GerminationeditorveterinaryworldNo ratings yet
- Estrogen DR DeanDocument8 pagesEstrogen DR DeanMochammad Halim NNo ratings yet
- LetrozoleDocument10 pagesLetrozoleThunnisa SivNo ratings yet
- 4925-300 E2 AccuBind ELISA Rev 4Document2 pages4925-300 E2 AccuBind ELISA Rev 4Roxana RoxNo ratings yet
- 2010 - Sex Differences in The Brain, Behavior, and Neuropsychiatric DisordersDocument16 pages2010 - Sex Differences in The Brain, Behavior, and Neuropsychiatric DisordersDiego Alejandro Londoño CorreaNo ratings yet
- Effects of Hormones and Hormone Therapy On Breast Tissue in Transgender Patients: A Concise ReviewDocument10 pagesEffects of Hormones and Hormone Therapy On Breast Tissue in Transgender Patients: A Concise ReviewperezhuertalexieNo ratings yet
- Testosterone: Mechanism of ActionDocument3 pagesTestosterone: Mechanism of Actionvasu VermaNo ratings yet
- Grade 10 - Quarter 3 Notes (All Subjects)Document44 pagesGrade 10 - Quarter 3 Notes (All Subjects)Rianne MoralesNo ratings yet
- Anabolism and CatabolismDocument8 pagesAnabolism and CatabolismEd RibeiroNo ratings yet
- Biosintesis Dan Mekanisme Kerja Hormon Repro Wanita PDFDocument39 pagesBiosintesis Dan Mekanisme Kerja Hormon Repro Wanita PDFsallynorcelinaNo ratings yet
- Fabre-Nys - 2007 - Sexual Behaviour in Ewes and Other RuminantsDocument8 pagesFabre-Nys - 2007 - Sexual Behaviour in Ewes and Other RuminantsGustavo Campos Bueno DiasNo ratings yet
- Vi45645ontfgd Dfger5344Document74 pagesVi45645ontfgd Dfger5344Hugo MendesNo ratings yet
- Puberty Blocking Medications: An OverviewDocument10 pagesPuberty Blocking Medications: An OverviewCata LinaNo ratings yet
- Harmacology of Strogens AND Estagens: H. KuhlDocument19 pagesHarmacology of Strogens AND Estagens: H. KuhlBruno de OliveiraNo ratings yet
- EndomeTRIO Manual USA and Canada English VersionDocument43 pagesEndomeTRIO Manual USA and Canada English Versionantik8No ratings yet
- BenzophenoneDocument20 pagesBenzophenoneYuuki93No ratings yet
- OBGYN End of Rotation Exam OutlineDocument35 pagesOBGYN End of Rotation Exam OutlinehevinpatelNo ratings yet