You might also like
- A Birds Eye View of The Aimst Uni (7 March 2015)Document174 pagesA Birds Eye View of The Aimst Uni (7 March 2015)M.Ravichandran100% (1)
- Seeing Eye To Eye: Vertebrate Eye Evolution and Adaptive RadiationDocument7 pagesSeeing Eye To Eye: Vertebrate Eye Evolution and Adaptive RadiationAprille Octaviano-AsiloNo ratings yet
- Penguin Skull PDFDocument15 pagesPenguin Skull PDFmedeNo ratings yet
- Exhibit PDFDocument15 pagesExhibit PDFJaffer RazaNo ratings yet
- 27 ArthropodsDocument63 pages27 Arthropodsapi-244168124No ratings yet
- Anatomy and Development of Oral Cavity and PharynxDocument14 pagesAnatomy and Development of Oral Cavity and PharynxGabriela MacoveanuNo ratings yet
- Projektarbeit Livia Pian2016-17 InglesDocument46 pagesProjektarbeit Livia Pian2016-17 InglesmariaNo ratings yet
- How To Tune Your MindDocument3 pagesHow To Tune Your MindSuhas DkNo ratings yet
- Astrology, A Free Online Course: AriesDocument23 pagesAstrology, A Free Online Course: Ariesnikos nevrosNo ratings yet
- Cosmic Freedom: David MolineauxDocument2 pagesCosmic Freedom: David Molineauxsalomon46No ratings yet
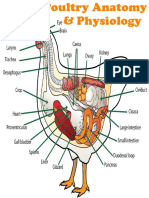
- PoultryAnatomyEng WDocument1 pagePoultryAnatomyEng WBryan NicollNo ratings yet
- ANATOMY OF PIGEON (Columba Domestica)Document12 pagesANATOMY OF PIGEON (Columba Domestica)Mellya RizkiNo ratings yet
- Common OstrichDocument165 pagesCommon OstrichHarish BhatNo ratings yet
- Matti Pitkanen - The Notion of Wave-Genome and DNA As Topological Quantum ComputerDocument15 pagesMatti Pitkanen - The Notion of Wave-Genome and DNA As Topological Quantum ComputerJoaokaaNo ratings yet
- Time - Internet Encyclopedia of PhilosophyDocument67 pagesTime - Internet Encyclopedia of Philosophyagf1001No ratings yet
- GasaitéarDocument200 pagesGasaitéarPMNo ratings yet
- © 2017 5th Dimensional Quantum Healing & Awareness by Author: Roisin HerreraDocument1 page© 2017 5th Dimensional Quantum Healing & Awareness by Author: Roisin HerrerasdaNo ratings yet
- The'Glass'House:' Crucible'of'Biodynamic'Agriculture': John - PaullDocument6 pagesThe'Glass'House:' Crucible'of'Biodynamic'Agriculture': John - PaullmahepoNo ratings yet
- Effect of Sucrose On HumansDocument19 pagesEffect of Sucrose On HumansSabashadoNo ratings yet
- The New Dialogue With The Spirit of The EarthDocument58 pagesThe New Dialogue With The Spirit of The EarthGucciGrumpsNo ratings yet
- The Three Logoi 4Document6 pagesThe Three Logoi 4masterascendedNo ratings yet
- Origins of Life: A D D S C DDocument23 pagesOrigins of Life: A D D S C DjonasgoNo ratings yet
- Foreword Arcana of LightDocument4 pagesForeword Arcana of LightNestor David MoralesNo ratings yet
- Endtime Prophecy That No Longer Applies To LuciferDocument14 pagesEndtime Prophecy That No Longer Applies To LuciferArnulfo Yu Laniba100% (1)
- 3 L 3 M 3 NtalDocument40 pages3 L 3 M 3 NtalBenjamminNo ratings yet
- Chapter 1 Introduction To Biology Lesson 1.1 and 1.7Document140 pagesChapter 1 Introduction To Biology Lesson 1.1 and 1.7Zetroc Jess100% (1)
- Methods of Studying Animal BehaviorDocument21 pagesMethods of Studying Animal BehaviorAntesh100% (1)
- SteinerDocument28 pagesSteinerSrijani BiswasNo ratings yet
- 24H Education PDFDocument28 pages24H Education PDFioanaaa_imNo ratings yet
- SS7R Study 1Document37 pagesSS7R Study 1DKelshawNo ratings yet
- Beginnings: 1.1 (Sorta) How Math WorksDocument11 pagesBeginnings: 1.1 (Sorta) How Math WorksBace HickamNo ratings yet
- Cephalopod - WikipediaDocument172 pagesCephalopod - WikipediaReynaLord Rafael-sanNo ratings yet
- Annie Cahn Fung - Paul Brunton, A Bridge Between India and The WestDocument83 pagesAnnie Cahn Fung - Paul Brunton, A Bridge Between India and The WestShabd_MysticNo ratings yet
- 1 2019 Herpetology PPTDocument45 pages1 2019 Herpetology PPTBryceNo ratings yet
- Nonvisual Photoreceptors of The PinealDocument36 pagesNonvisual Photoreceptors of The Pinealrambo_style19No ratings yet
- Plant Cell and Animal CellDocument8 pagesPlant Cell and Animal CelltipsyleopardNo ratings yet
- TICHENOR Theory of Reincarnation PDFDocument38 pagesTICHENOR Theory of Reincarnation PDFrobertfirth2759No ratings yet
- A Bird's Eye View of The Aimst UniversityDocument199 pagesA Bird's Eye View of The Aimst UniversitySubhash Janardhan Bhore, PhDNo ratings yet
- The Calendar of The SoulDocument1 pageThe Calendar of The Soul3adrianNo ratings yet
- A History of EntomologicalDocument20 pagesA History of EntomologicalBarzola HerlissNo ratings yet
- The European Eel (Anguilla Anguilla, Linnaeus), Its Lifecycle, Evolution and Reproduction: A Literature ReviewDocument33 pagesThe European Eel (Anguilla Anguilla, Linnaeus), Its Lifecycle, Evolution and Reproduction: A Literature ReviewVikrant GaikwadNo ratings yet
- Civilization Book4Document23 pagesCivilization Book4api-176452956No ratings yet
- Insect MorphologyDocument10 pagesInsect MorphologyDragan GrčakNo ratings yet
- Snakes On An Enrichment Programme?!: Shelby Stanfield, MSC James Brereton, MSCDocument11 pagesSnakes On An Enrichment Programme?!: Shelby Stanfield, MSC James Brereton, MSCWarner LlaveNo ratings yet
- ZarathustraDocument13 pagesZarathustraEtra PublishingNo ratings yet
- World Literature Greek MythDocument1 pageWorld Literature Greek Mythbulkz007No ratings yet
- 4 MyologyDocument57 pages4 MyologyAbhijith S. PNo ratings yet
- History of MathematicsDocument5 pagesHistory of MathematicsAsley Kate BienNo ratings yet
- LeadBeater Service of Healing 02Document5 pagesLeadBeater Service of Healing 02ahndres12No ratings yet
- Star ScholarDocument16 pagesStar ScholarMatthew SaintNo ratings yet
- Advanced ThermodynamicsDocument6 pagesAdvanced ThermodynamicsKumar Kote100% (1)
- Class AvesDocument11 pagesClass AveshemavarshiniNo ratings yet
- General Characters of Class AvesDocument10 pagesGeneral Characters of Class AvesGEETA MOHAN100% (3)
- Chapter 1: Introduction To Biology 1.1the Study of BiologyDocument3 pagesChapter 1: Introduction To Biology 1.1the Study of BiologyCikguAgnesNo ratings yet
- Thermo RegulationDocument12 pagesThermo RegulationSnapeSnapeNo ratings yet
- Adaptation of Art Education To Suit The Needs of Young People in MalaysiaDocument15 pagesAdaptation of Art Education To Suit The Needs of Young People in MalaysiaAzlan Andi100% (1)
- The Four CausesDocument27 pagesThe Four CausesVia Suci RohmahNo ratings yet
- Eastern PhilosophyDocument70 pagesEastern PhilosophyRedimto RoseteNo ratings yet
- Handbook of Bird Biology - CH 4Document4 pagesHandbook of Bird Biology - CH 4butylacetateNo ratings yet
- Quiz Organ SysDocument1 pageQuiz Organ SysMelva GuerraNo ratings yet
- The Design of The Avian Respiratory System: Development and Morphology - A Review A.D.FirdousDocument24 pagesThe Design of The Avian Respiratory System: Development and Morphology - A Review A.D.FirdousIndian Journal of Veterinary and Animal Sciences RNo ratings yet
- Aquila Pomarina X Clanga ClangaDocument14 pagesAquila Pomarina X Clanga ClangaCristinaNo ratings yet
- VertebratesDocument63 pagesVertebratesgenusxyz100% (1)
- Class AvesDocument4 pagesClass AvesREYGILINE CLEMENTENo ratings yet
- Cap. 06 - Avian Anatomy and PhysiologyDocument65 pagesCap. 06 - Avian Anatomy and PhysiologyNailson JúniorNo ratings yet
- Class AvesDocument12 pagesClass AvesTani SinghNo ratings yet
- Types of FeatherDocument7 pagesTypes of Featherlooperrr05No ratings yet
- Science RespuestasDocument18 pagesScience RespuestasNancy LuNo ratings yet
- Poultry ProductionDocument16 pagesPoultry ProductionAthena NocetoNo ratings yet
- Build A Bird: Bird Adaptions: Teacher/Volunteer Summary Adapted From Shorebird Sister School, "To Build A Shorebird"Document5 pagesBuild A Bird: Bird Adaptions: Teacher/Volunteer Summary Adapted From Shorebird Sister School, "To Build A Shorebird"Shushovan Usha BanikNo ratings yet
- Group 1 - Chicken Dissection PDFDocument37 pagesGroup 1 - Chicken Dissection PDFMohammad Omar BacaramanNo ratings yet
- 4th Q Module GEN BIO 2 Week 5 6Document19 pages4th Q Module GEN BIO 2 Week 5 6Hannah CastroNo ratings yet
- Animal Science - Narrative ReportDocument10 pagesAnimal Science - Narrative ReportRowena Kig-isan MeloyNo ratings yet
- Avian Skeletal System - EXtensionDocument3 pagesAvian Skeletal System - EXtensionNagyeongNo ratings yet
- Poultry Health and Management 359p Color Picture PDFDocument359 pagesPoultry Health and Management 359p Color Picture PDFRaj Ghankute RGNo ratings yet
- AVIAN Respiratory SystemDocument7 pagesAVIAN Respiratory Systemtaner_soysurenNo ratings yet
- Chapter 9 Respiratory Systems NotesDocument11 pagesChapter 9 Respiratory Systems NotesCaleb DodgeNo ratings yet
- Diseases Poultry Diagnosis SymptomsDocument15 pagesDiseases Poultry Diagnosis SymptomsTufail Banday100% (1)
- Conditions in Common Avian and Aquatic SpeciesDocument34 pagesConditions in Common Avian and Aquatic SpeciesŠhâh NawazNo ratings yet
- H SC GlossaryDocument64 pagesH SC Glossaryjose cabelloNo ratings yet
- 6 Avian Medicine Principles and Application - HarrisonDocument23 pages6 Avian Medicine Principles and Application - Harrisonmaluka09No ratings yet
- Chicken Anatomy & PhysiologyDocument26 pagesChicken Anatomy & PhysiologySanjeev ChaudharyNo ratings yet
- 4H Dog Skeletal AnatomyDocument2 pages4H Dog Skeletal Anatomybhanny2No ratings yet
- Avian Skeletal System - Robert Porter PDFDocument16 pagesAvian Skeletal System - Robert Porter PDFJohn MillerNo ratings yet
- Emergency RaptorsDocument24 pagesEmergency RaptorsjudithNo ratings yet
- Librito ExóticosDocument92 pagesLibrito ExóticosLuis GonzalezNo ratings yet
- Veterinary Anatomy Coloring Book - Anatomy AcademyDocument54 pagesVeterinary Anatomy Coloring Book - Anatomy AcademyJos torresNo ratings yet
- Classification: Down FeathersDocument2 pagesClassification: Down FeathersCarlo QuinlogNo ratings yet
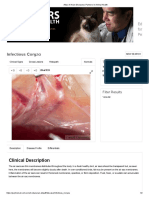
- Atlas of Avian Diseases - Partners in Animal HealthDocument2 pagesAtlas of Avian Diseases - Partners in Animal HealthShah NawazNo ratings yet