You might also like
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (121)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (400)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5795)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1091)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (345)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Learn C++ Programming LanguageDocument322 pagesLearn C++ Programming LanguageAli Eb100% (8)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- HiPerMat 2012 KasselDocument1,059 pagesHiPerMat 2012 KasselAna Mafalda MatosNo ratings yet
- Critical Point of View: A Wikipedia ReaderDocument195 pagesCritical Point of View: A Wikipedia Readerrll307No ratings yet
- Internship Report On Service IndustryDocument50 pagesInternship Report On Service Industrybbaahmad89No ratings yet
- SLC 10 ScienceDocument16 pagesSLC 10 SciencePffflyers KurnawanNo ratings yet
- Free and Forced Vortex PDFDocument4 pagesFree and Forced Vortex PDFRana HanzlaNo ratings yet
- 1 SMDocument9 pages1 SMSelly ManaluNo ratings yet
- Joshi C M, Vyas Y - Extensions of Certain Classical Integrals of Erdélyi For Gauss Hypergeometric Functions - J. Comput. and Appl. Mat. 160 (2003) 125-138Document14 pagesJoshi C M, Vyas Y - Extensions of Certain Classical Integrals of Erdélyi For Gauss Hypergeometric Functions - J. Comput. and Appl. Mat. 160 (2003) 125-138Renee BravoNo ratings yet
- PERFORM Toolkit 3 1 Release Notes 1.0 OnlinePDFDocument18 pagesPERFORM Toolkit 3 1 Release Notes 1.0 OnlinePDFJose HugoNo ratings yet
- Solarbotics Wheel Watcher Encoder ManualDocument10 pagesSolarbotics Wheel Watcher Encoder ManualYash SharmaNo ratings yet
- Practical Means For Energy-Based Analyses of Disproportionate Collapse PotentialDocument13 pagesPractical Means For Energy-Based Analyses of Disproportionate Collapse PotentialNhut NguyenNo ratings yet
- Kelompok 5Document16 pagesKelompok 5bintangNo ratings yet
- Property PalleteDocument5 pagesProperty PalletePranay PatelNo ratings yet
- Charity Connecting System: M.Archana, K.MouthamiDocument5 pagesCharity Connecting System: M.Archana, K.MouthamiShainara Dela TorreNo ratings yet
- 2.2 Lesson Notes FA12Document24 pages2.2 Lesson Notes FA12JAPNo ratings yet
- EnglishFile4e Intermediate Plus TG PCM Vocab 1BDocument1 pageEnglishFile4e Intermediate Plus TG PCM Vocab 1Bayşe adaliNo ratings yet
- D027B OmronDocument141 pagesD027B OmronirfanWPKNo ratings yet
- Raglio 2015 Effects of Music and Music Therapy On Mood in Neurological Patients PDFDocument12 pagesRaglio 2015 Effects of Music and Music Therapy On Mood in Neurological Patients PDFsimaso0% (1)
- Meeting Wise Agenda TemplateDocument1 pageMeeting Wise Agenda Templateapi-416923934No ratings yet
- For Purposeful and Meaningful Technology Integration in The ClassroomDocument19 pagesFor Purposeful and Meaningful Technology Integration in The ClassroomVincent Chris Ellis TumaleNo ratings yet
- #Stopableism: Reduksi Stigma Kepada Penyandang Disabilitas Melalui Intervensi Bias ImplisitDocument15 pages#Stopableism: Reduksi Stigma Kepada Penyandang Disabilitas Melalui Intervensi Bias ImplisitDennis SuhardiniNo ratings yet
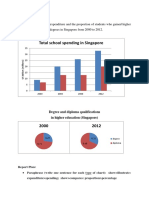
- Total School Spending in Singapore: Task 1 020219Document6 pagesTotal School Spending in Singapore: Task 1 020219viper_4554No ratings yet
- Barry Farm Powerpoint SlidesDocument33 pagesBarry Farm Powerpoint SlidessarahNo ratings yet
- Evaluation FormDocument3 pagesEvaluation FormFreyjaa MabelinNo ratings yet
- Role of Wickability On The Critical Heat Flux of Structured Superhydrophilic SurfacesDocument10 pagesRole of Wickability On The Critical Heat Flux of Structured Superhydrophilic Surfacesavi0341No ratings yet
- Affective DomainDocument3 pagesAffective DomainJm Enriquez Dela Cruz50% (2)
- Contoh RefleksiDocument7 pagesContoh RefleksiIzzat HanifNo ratings yet
- Spirolab IIIDocument1 pageSpirolab IIIDzenan DelijaNo ratings yet
- Mutants & Masterminds 1st Ed Character Generater (Xls Spreadsheet)Document89 pagesMutants & Masterminds 1st Ed Character Generater (Xls Spreadsheet)WesleyNo ratings yet
- Report Swtich Cisco Pass DNSDocument91 pagesReport Swtich Cisco Pass DNSDenis Syst Laime LópezNo ratings yet