You might also like
- Nutrition Conference for Feed Manufacturers: University of Nottingham, Volume 7From EverandNutrition Conference for Feed Manufacturers: University of Nottingham, Volume 7No ratings yet
- tmpAC TMPDocument12 pagestmpAC TMPFrontiersNo ratings yet
- Cowling Et Al-2020-Scientific ReportsDocument8 pagesCowling Et Al-2020-Scientific ReportsMark Christian DiamanteNo ratings yet
- To Mendel's Laws. These Traits Can Also Be Affected by The Environment To Varying DegreesDocument3 pagesTo Mendel's Laws. These Traits Can Also Be Affected by The Environment To Varying DegreesSunil BhavsarNo ratings yet
- Genetic Divergence Among Maize (Zea Mays L.) Inbred Lines Using Morphometric MarkersDocument15 pagesGenetic Divergence Among Maize (Zea Mays L.) Inbred Lines Using Morphometric MarkersIJAR JOURNALNo ratings yet
- Transcriptome Pro Ling of Two Hybrid Rice Provides Insights Into The Genetic Basis of HeterosisDocument20 pagesTranscriptome Pro Ling of Two Hybrid Rice Provides Insights Into The Genetic Basis of HeterosisAl 015No ratings yet
- 32 Ijasroct201732Document6 pages32 Ijasroct201732TJPRC PublicationsNo ratings yet
- Nachimuthu Article AnalysisOfPopulationStructureADocument25 pagesNachimuthu Article AnalysisOfPopulationStructureAhannaNo ratings yet
- Agronomy 11 02528 v2Document18 pagesAgronomy 11 02528 v2Ali SherNo ratings yet
- 1 s2.0 S1674205218303642 MainDocument3 pages1 s2.0 S1674205218303642 MainRodrigo RuizNo ratings yet
- Genotyping and Phenotyping Strategies For Genetic Improvement of Meat Quality and Carcass Composition in SwineDocument16 pagesGenotyping and Phenotyping Strategies For Genetic Improvement of Meat Quality and Carcass Composition in Swinestephano portugalNo ratings yet
- C1107021636 PDFDocument21 pagesC1107021636 PDFTeflon SlimNo ratings yet
- Allelic Analysis of Sheath Blight Resistance With Association Mapping in RiceDocument10 pagesAllelic Analysis of Sheath Blight Resistance With Association Mapping in Rice10sgNo ratings yet
- Fasahat Et Al., 2016Document25 pagesFasahat Et Al., 2016Edjane FreitasNo ratings yet
- Principles and Utilization of Combining Ability in PDFDocument25 pagesPrinciples and Utilization of Combining Ability in PDFEdjane FreitasNo ratings yet
- Plant Biotechnology Journal - 2019 - Abbai - Haplotype Analysis of Key Genes Governing Grain Yield and Quality TraitsDocument11 pagesPlant Biotechnology Journal - 2019 - Abbai - Haplotype Analysis of Key Genes Governing Grain Yield and Quality TraitsMay Htet AungNo ratings yet
- Plantgenome2017 09 0085 PDFDocument21 pagesPlantgenome2017 09 0085 PDFJawaria IshfaqNo ratings yet
- Study On Combining Ability and Heterosis in Maize (Zea Mays L.) Using Partial Diallel AnalysisDocument7 pagesStudy On Combining Ability and Heterosis in Maize (Zea Mays L.) Using Partial Diallel AnalysisPremier PublishersNo ratings yet
- Yu Et Al., 2021Document11 pagesYu Et Al., 2021Hercules SantosNo ratings yet
- Breeding Self-Pollinated PlantsDocument7 pagesBreeding Self-Pollinated PlantsAdis MogesNo ratings yet
- Horticulturae 10 00120Document16 pagesHorticulturae 10 00120omaragronomieNo ratings yet
- Heterosis For Seed Yield and Yield Contributing Characters of Kabuli X Kabuli Crosses of Chickpea (Cicer Arietinum L.)Document8 pagesHeterosis For Seed Yield and Yield Contributing Characters of Kabuli X Kabuli Crosses of Chickpea (Cicer Arietinum L.)Pallavi SasaneNo ratings yet
- 1 s2.0 S0044848622003453 MainDocument8 pages1 s2.0 S0044848622003453 MainRama BangeraNo ratings yet
- Jurnal 1Document7 pagesJurnal 1Muhammad Maulana SidikNo ratings yet
- Variation, Correlation and Path-Coefficient Study in Groundnut Breeding Lines of OdishaDocument12 pagesVariation, Correlation and Path-Coefficient Study in Groundnut Breeding Lines of OdishaTJPRC PublicationsNo ratings yet
- Assessment of Breeding Programs Sustainability: Application of Phenotypic and Genomic Indicators To A North European Grain Maize ProgramDocument14 pagesAssessment of Breeding Programs Sustainability: Application of Phenotypic and Genomic Indicators To A North European Grain Maize Programcata2099No ratings yet
- Taxonomic and Functional Assessment Using Metatranscriptomics Reveals The Effect of Angus Cattle On Rumen Microbial SignaturesDocument15 pagesTaxonomic and Functional Assessment Using Metatranscriptomics Reveals The Effect of Angus Cattle On Rumen Microbial SignaturesAndré Luis NevesNo ratings yet
- PNAS 1997 Yu 9226 31Document6 pagesPNAS 1997 Yu 9226 31Dani Pazmiño BonillaNo ratings yet
- Combining Pure-Line and Cross-Bred Information in Poultry BreedingDocument6 pagesCombining Pure-Line and Cross-Bred Information in Poultry BreedingRvk NairNo ratings yet
- Genome Editing Livestock Breeding ProgramsDocument17 pagesGenome Editing Livestock Breeding Programsmirielys.rondaNo ratings yet
- Co-So-Di-Truyen-Chon-Giong-Cay-Trong - Agronomy-10-00180 - (Cuuduongthancong - Com)Document15 pagesCo-So-Di-Truyen-Chon-Giong-Cay-Trong - Agronomy-10-00180 - (Cuuduongthancong - Com)Hai HeoNo ratings yet
- Genetic Analyses of Wheat and Molecular Marker-Assisted Breeding, Volume 1Document550 pagesGenetic Analyses of Wheat and Molecular Marker-Assisted Breeding, Volume 1LeoNo ratings yet
- HTPdb and HTPtools-正式发表版Document10 pagesHTPdb and HTPtools-正式发表版Holibut HoNo ratings yet
- Molecular Versus Morphological Markers To Describe Variability in Sugar Cane (Saccharum Officinarum) For Germplasm Management and ConservationDocument21 pagesMolecular Versus Morphological Markers To Describe Variability in Sugar Cane (Saccharum Officinarum) For Germplasm Management and ConservationBlaxez YTNo ratings yet
- Genetic Evaluation of Tropical Popcorn Inbred Lines Using BLUPDocument10 pagesGenetic Evaluation of Tropical Popcorn Inbred Lines Using BLUPKlayton Flávio MilaniNo ratings yet
- Rice Genomics ResearchDocument27 pagesRice Genomics ResearchDiane Fernandez BenesioNo ratings yet
- Barley PaperDocument5 pagesBarley PaperNeus Prieto DiezNo ratings yet
- Multi-Breed Genomic Predictions and Functional Variants For Fertility of Tropical BullsDocument19 pagesMulti-Breed Genomic Predictions and Functional Variants For Fertility of Tropical BullsBahlibiNo ratings yet
- Genetic Associations, Variability and Diversity in Biochemical and Morphological Seed Characters in Indian Grass Pea (Lathyrussativus L.) AccessionsDocument7 pagesGenetic Associations, Variability and Diversity in Biochemical and Morphological Seed Characters in Indian Grass Pea (Lathyrussativus L.) AccessionsHappey Flower GeneralNo ratings yet
- Mas 4Document26 pagesMas 4Nofiya RuswantiNo ratings yet
- GWAS IntroductionDocument4 pagesGWAS IntroductionTouqeer AhmadNo ratings yet
- Genome-Wide Association Mapping: A Case Study in Bread Wheat (Triticum Aestivum L.)Document22 pagesGenome-Wide Association Mapping: A Case Study in Bread Wheat (Triticum Aestivum L.)Ahmad MasoodNo ratings yet
- Of Trzbolzum Castaneum: HazelDocument22 pagesOf Trzbolzum Castaneum: HazelZask CrushNo ratings yet
- Study of Genetic Variability and Character Association For Yield and Yield Related Traits in f3 Generation of Blackgram (Vignamungo (L.) Hepper)Document13 pagesStudy of Genetic Variability and Character Association For Yield and Yield Related Traits in f3 Generation of Blackgram (Vignamungo (L.) Hepper)Mamta AgarwalNo ratings yet
- tmp8FD0 TMPDocument10 pagestmp8FD0 TMPFrontiersNo ratings yet
- Study of Cgms Based Pigeonpea (Cajanus Cajan (L.) Millsp) Hybrids in Terms of Combining AbilityDocument6 pagesStudy of Cgms Based Pigeonpea (Cajanus Cajan (L.) Millsp) Hybrids in Terms of Combining AbilityTJPRC PublicationsNo ratings yet
- Diallel Genetic Analysis For Multiple Traits in EgDocument20 pagesDiallel Genetic Analysis For Multiple Traits in EgAde MulyanaNo ratings yet
- 19 Maize Trait Association 2022Document5 pages19 Maize Trait Association 2022Bandhan ThapaNo ratings yet
- Genetic Diversity of A Maize Association Population With Restricted PhenologyDocument12 pagesGenetic Diversity of A Maize Association Population With Restricted PhenologyFrontiersNo ratings yet
- Cacahuate Genetic Analysis of YieldDocument5 pagesCacahuate Genetic Analysis of YieldVictorNo ratings yet
- Heterotic PoolsDocument19 pagesHeterotic PoolsVinaykumarNo ratings yet
- 1 s2.0 S1751731118001489 MainDocument15 pages1 s2.0 S1751731118001489 MainMaha RanuNo ratings yet
- Wheat DroughtDocument8 pagesWheat DroughtInes Jlassi Ep MabroukNo ratings yet
- Heterotic Potential, Potence Ratio, Combining Ability and Genetic Control of Seed Vigour Traits For Yield Improvement in Cucumber (Cucumis Sativus)Document8 pagesHeterotic Potential, Potence Ratio, Combining Ability and Genetic Control of Seed Vigour Traits For Yield Improvement in Cucumber (Cucumis Sativus)Dr Sandeep KumarNo ratings yet
- Thesis On Hybrid RiceDocument7 pagesThesis On Hybrid Riceazyxppzcf100% (2)
- AndyGWAS-RevisedManuscript FileDocument26 pagesAndyGWAS-RevisedManuscript FileZeleke WondimuNo ratings yet
- Common Bean Genomics and Its Applications in Breeding ProgramsDocument22 pagesCommon Bean Genomics and Its Applications in Breeding ProgramsrmbichiNo ratings yet
- Genome Wide Association Studies and Genomic Selection - Promises and PremisesDocument16 pagesGenome Wide Association Studies and Genomic Selection - Promises and Premisessalman672003No ratings yet
- Selection of Core Collections From Kartiksail and Dhaliboro Rice Landraces of BangladeshDocument11 pagesSelection of Core Collections From Kartiksail and Dhaliboro Rice Landraces of BangladeshMd Ashikur RahmanNo ratings yet
- Database of Cattle Candidate Genes and Genetic MarkersDocument20 pagesDatabase of Cattle Candidate Genes and Genetic MarkersrohishaakNo ratings yet
- Conc 0.0 0.1 0.2 0.3 100W TN FTN PH STH SPL FLL AWL SPN SPS SPW STW PLWDocument1 pageConc 0.0 0.1 0.2 0.3 100W TN FTN PH STH SPL FLL AWL SPN SPS SPW STW PLW53melmelNo ratings yet
- SNPBC Proceedings PDFDocument376 pagesSNPBC Proceedings PDF53melmelNo ratings yet
- Genotype Environment: DTDPDM DD DRDocument1 pageGenotype Environment: DTDPDM DD DR53melmelNo ratings yet
- SNPBC Proceedings PDFDocument376 pagesSNPBC Proceedings PDF53melmelNo ratings yet
- SNPBC Proceedings PDFDocument376 pagesSNPBC Proceedings PDF53melmelNo ratings yet
- Hort Sci-Sample Paper PDFDocument7 pagesHort Sci-Sample Paper PDF53melmelNo ratings yet
- Fourier Blobs: Subwrld2 Subwrld2Document2 pagesFourier Blobs: Subwrld2 Subwrld253melmelNo ratings yet
- DsagfDocument5 pagesDsagf53melmelNo ratings yet
- Answers Part 2Document64 pagesAnswers Part 2Rynjeff Lui-Pio100% (1)
- Diet & Nutrition Advanced Ver 1Document52 pagesDiet & Nutrition Advanced Ver 1Ina100% (1)
- 21 Benign Skin TumorsDocument4 pages21 Benign Skin TumorsAbdul Ghaffar AbdullahNo ratings yet
- Inflammation. Etiology. Vascular Changes. Cellular Events in Inflammation. Acute Inflammation. Morphologic PatternsDocument57 pagesInflammation. Etiology. Vascular Changes. Cellular Events in Inflammation. Acute Inflammation. Morphologic PatternsZauzaNo ratings yet
- 2 Thrombosed HemorrhoidsDocument5 pages2 Thrombosed HemorrhoidsMarko LovrićNo ratings yet
- Digestive System Charts 1Document14 pagesDigestive System Charts 1api-306762527No ratings yet
- 02 - Recalibrating Gavi 5.0 in Light of COVID-19 and Successful Replenis..Document19 pages02 - Recalibrating Gavi 5.0 in Light of COVID-19 and Successful Replenis..aman_sasanNo ratings yet
- Genetic Engineering PDFDocument50 pagesGenetic Engineering PDFBrian Rey L. AbingNo ratings yet
- Bonus BooksDocument540 pagesBonus BooksDanielle Moore100% (2)
- ForrestDocument4 pagesForrestAbulHasan Idrus IstarNo ratings yet
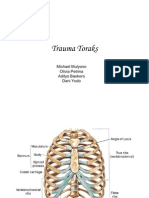
- Trauma ToraksDocument30 pagesTrauma ToraksMichael KwanNo ratings yet
- Beta Blocker TitrationDocument2 pagesBeta Blocker TitrationAwalunisa IstiqomahNo ratings yet
- BobackDocument4 pagesBobackapi-324391171No ratings yet
- Radiologi Intervensi Di IndiaDocument3 pagesRadiologi Intervensi Di IndiaPutu Adi SusantaNo ratings yet
- 4biomechanics of Removable Partial DenturesDocument15 pages4biomechanics of Removable Partial DenturesDemilyadioppy AbevitNo ratings yet
- Behavior and Social SciencesDocument10 pagesBehavior and Social SciencesRoxana Alexandra BogosNo ratings yet
- Tricks To Remember Important Days-All-MonthsDocument2 pagesTricks To Remember Important Days-All-MonthsSenthikumar MuraliNo ratings yet
- Auras PDFDocument24 pagesAuras PDFvijayant pandeyNo ratings yet
- Role of CT Scan in Diagnosis of Acute AppendicitisDocument6 pagesRole of CT Scan in Diagnosis of Acute AppendicitisIndahNo ratings yet
- VITAMINS-Carol Jane Lorenzo, RMTDocument8 pagesVITAMINS-Carol Jane Lorenzo, RMTREYES, JAN MERCK M.No ratings yet
- Drugs-1.ppt 0Document22 pagesDrugs-1.ppt 0Esraa BahaaNo ratings yet
- Revalida 1Document3 pagesRevalida 1herrabiel solisNo ratings yet
- Script For OrientationDocument2 pagesScript For OrientationAngela CudiaNo ratings yet
- Review - Spinal Cord Injury - 2Document1 pageReview - Spinal Cord Injury - 2Sarah MendozaNo ratings yet
- Report Text LatihanDocument12 pagesReport Text Latihanbenta pasyahNo ratings yet
- Danger of Smoking in The MorningDocument2 pagesDanger of Smoking in The MorningAndi Intan AuliaNo ratings yet
- 3b Oral Exam Practice - Mock Exam THREE - ANSWER Guide PDFDocument7 pages3b Oral Exam Practice - Mock Exam THREE - ANSWER Guide PDFJyothsna SridharNo ratings yet
- Primal Essence - A Man's Guide To Maximizing Testosterone NaturallyDocument46 pagesPrimal Essence - A Man's Guide To Maximizing Testosterone NaturallyKristiyan GrigorovNo ratings yet
- Dermatology - James G. Marks, Jeffrey J. Miller - Lookingbill and Marks' Principles of Dermatology (2018, Elsevier) (Dragged)Document9 pagesDermatology - James G. Marks, Jeffrey J. Miller - Lookingbill and Marks' Principles of Dermatology (2018, Elsevier) (Dragged)nh2411No ratings yet
- A Stepwise Enteral Nutrition Algorithm For Critically (Susan Dan Nilesh Mehta)Document7 pagesA Stepwise Enteral Nutrition Algorithm For Critically (Susan Dan Nilesh Mehta)NyomanGinaHennyKristiantiNo ratings yet
- All That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesFrom EverandAll That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesRating: 4.5 out of 5 stars4.5/5 (397)
- Gut: the new and revised Sunday Times bestsellerFrom EverandGut: the new and revised Sunday Times bestsellerRating: 4 out of 5 stars4/5 (393)
- Why We Die: The New Science of Aging and the Quest for ImmortalityFrom EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityRating: 4 out of 5 stars4/5 (5)
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceFrom EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceRating: 4.5 out of 5 stars4.5/5 (517)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisFrom EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisRating: 3.5 out of 5 stars3.5/5 (2)
- 10% Human: How Your Body's Microbes Hold the Key to Health and HappinessFrom Everand10% Human: How Your Body's Microbes Hold the Key to Health and HappinessRating: 4 out of 5 stars4/5 (33)
- Tales from Both Sides of the Brain: A Life in NeuroscienceFrom EverandTales from Both Sides of the Brain: A Life in NeuroscienceRating: 3 out of 5 stars3/5 (18)
- A Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsFrom EverandA Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsRating: 4.5 out of 5 stars4.5/5 (6)
- The Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionFrom EverandThe Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionRating: 4 out of 5 stars4/5 (812)
- Return of the God Hypothesis: Three Scientific Discoveries That Reveal the Mind Behind the UniverseFrom EverandReturn of the God Hypothesis: Three Scientific Discoveries That Reveal the Mind Behind the UniverseRating: 4.5 out of 5 stars4.5/5 (52)
- Fast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperFrom EverandFast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperRating: 4.5 out of 5 stars4.5/5 (15)
- Who's in Charge?: Free Will and the Science of the BrainFrom EverandWho's in Charge?: Free Will and the Science of the BrainRating: 4 out of 5 stars4/5 (65)
- Gut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)From EverandGut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)Rating: 4 out of 5 stars4/5 (411)
- Undeniable: How Biology Confirms Our Intuition That Life Is DesignedFrom EverandUndeniable: How Biology Confirms Our Intuition That Life Is DesignedRating: 4 out of 5 stars4/5 (11)
- Buddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomFrom EverandBuddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomRating: 4 out of 5 stars4/5 (216)
- Good Without God: What a Billion Nonreligious People Do BelieveFrom EverandGood Without God: What a Billion Nonreligious People Do BelieveRating: 4 out of 5 stars4/5 (66)
- The Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorFrom EverandThe Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorNo ratings yet
- Seven and a Half Lessons About the BrainFrom EverandSeven and a Half Lessons About the BrainRating: 4 out of 5 stars4/5 (110)
- Human: The Science Behind What Makes Your Brain UniqueFrom EverandHuman: The Science Behind What Makes Your Brain UniqueRating: 3.5 out of 5 stars3.5/5 (38)
- Moral Tribes: Emotion, Reason, and the Gap Between Us and ThemFrom EverandMoral Tribes: Emotion, Reason, and the Gap Between Us and ThemRating: 4.5 out of 5 stars4.5/5 (115)
- The Rise and Fall of the Dinosaurs: A New History of a Lost WorldFrom EverandThe Rise and Fall of the Dinosaurs: A New History of a Lost WorldRating: 4 out of 5 stars4/5 (595)
- Minds Make Societies: How Cognition Explains the World Humans CreateFrom EverandMinds Make Societies: How Cognition Explains the World Humans CreateRating: 4.5 out of 5 stars4.5/5 (23)
- A Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouFrom EverandA Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouRating: 4.5 out of 5 stars4.5/5 (62)
- The Invention of Tomorrow: A Natural History of ForesightFrom EverandThe Invention of Tomorrow: A Natural History of ForesightRating: 4.5 out of 5 stars4.5/5 (5)
- The Lives of Bees: The Untold Story of the Honey Bee in the WildFrom EverandThe Lives of Bees: The Untold Story of the Honey Bee in the WildRating: 4.5 out of 5 stars4.5/5 (44)