You might also like
- Long-Range Organization of Tandem Arrays of A Satellite DNA at The Centromeres of Human ChromosomesDocument5 pagesLong-Range Organization of Tandem Arrays of A Satellite DNA at The Centromeres of Human ChromosomesMariano HorganNo ratings yet
- 385 FullDocument8 pages385 FullEduardo MendozaNo ratings yet
- Kathleen Helm-Bychowski and Joel Cracraft "F: Mol. Bid. Evol. 1 O (6) : 1196-12 14. 1993Document19 pagesKathleen Helm-Bychowski and Joel Cracraft "F: Mol. Bid. Evol. 1 O (6) : 1196-12 14. 1993muffindidiNo ratings yet
- De Novo SATAC Formation 1 Chrom Res 1996Document14 pagesDe Novo SATAC Formation 1 Chrom Res 1996Robert KatonaNo ratings yet
- Hal 00893898Document31 pagesHal 00893898Vesee KavezeriNo ratings yet
- Physical Map Drosophila Melanogaster Divisions: Towards A of The GenomicDocument10 pagesPhysical Map Drosophila Melanogaster Divisions: Towards A of The GenomicJuvelin AuliaNo ratings yet
- BF 00314252Document2 pagesBF 00314252Lata DeshmukhNo ratings yet
- Pubmed Article of Crotalus HorridusDocument3 pagesPubmed Article of Crotalus HorridusPraveen ManoharNo ratings yet
- Nature VerdaderoDocument5 pagesNature VerdaderoCarolina RicárdezNo ratings yet
- Kitao 1996Document9 pagesKitao 1996M4shroomNo ratings yet
- Robe 2010 Amd DDCDocument12 pagesRobe 2010 Amd DDCGilberto Cavalheiro VieiraNo ratings yet
- The Caenorhabditis Elegans Schnurri Homolog Sma-9 Mediates Stage-And Cell Type-Specific Responses To DBL-1 BMP-related SignalingDocument12 pagesThe Caenorhabditis Elegans Schnurri Homolog Sma-9 Mediates Stage-And Cell Type-Specific Responses To DBL-1 BMP-related SignalingAziz RosyidNo ratings yet
- Check - Rapposa - Highly Conserved D-Loop Sequences in Woolly Mouse Opossums MarmosaDocument8 pagesCheck - Rapposa - Highly Conserved D-Loop Sequences in Woolly Mouse Opossums Marmosagbitencourt.bioNo ratings yet
- 1931 (2000) Toomas Kivisild and Richard Villems: ScienceDocument6 pages1931 (2000) Toomas Kivisild and Richard Villems: ScienceSrimeenakshi ShankarNo ratings yet
- Unit-Ii Asm 2022-23Document33 pagesUnit-Ii Asm 2022-23AnshnuNo ratings yet
- C Banding in Mus MusculusDocument10 pagesC Banding in Mus MusculusFlavio DávilaNo ratings yet
- Lande Diner2005Document7 pagesLande Diner2005Florencia FirenzeNo ratings yet
- Philippe Cluzel, Anne Lebrun, Christoph Heller, Richard Lavery, Jean-Louis Viovy, Didier Chatenay and François Caron - DNA: An Extensible MoleculeDocument4 pagesPhilippe Cluzel, Anne Lebrun, Christoph Heller, Richard Lavery, Jean-Louis Viovy, Didier Chatenay and François Caron - DNA: An Extensible MoleculeUylrikkNo ratings yet
- In Brief: Complexity On The NanoscaleDocument1 pageIn Brief: Complexity On The Nanoscaletopgun625No ratings yet
- Brown 1986Document9 pagesBrown 1986Liche Puello CaballeroNo ratings yet
- Monteiro Et Al., 2003 PDFDocument10 pagesMonteiro Et Al., 2003 PDFGutierrez Guarnizo SneiderNo ratings yet
- of 18 September 2013Document17 pagesof 18 September 2013Jean-claude PerezNo ratings yet
- A Genetic Analysis of The Cytological Region 46F-47B Containing The DrosophilaDocument7 pagesA Genetic Analysis of The Cytological Region 46F-47B Containing The DrosophilaeddubrovskyNo ratings yet
- The Origin of Human Chromosome 1 and Its Homologs in Placental MammalsDocument9 pagesThe Origin of Human Chromosome 1 and Its Homologs in Placental MammalspedrojcadNo ratings yet
- 11 ForensicsDocument29 pages11 ForensicsNithin JanardhananNo ratings yet
- Id of Native American Founder mtDNAsDocument13 pagesId of Native American Founder mtDNAsGerardo GonzalezNo ratings yet
- Connell Hofmann 2012Document5 pagesConnell Hofmann 2012AncaNo ratings yet
- STreptomyces Plasmid GiantDocument3 pagesSTreptomyces Plasmid GiantA2 ZNo ratings yet
- 1 s2.0 S1055790398906029 MainDocument10 pages1 s2.0 S1055790398906029 Mainjanaalharbi116No ratings yet
- (Li, W., Freundenberg, J.) Mappability and Read LengthDocument7 pages(Li, W., Freundenberg, J.) Mappability and Read LengthJohn SmithNo ratings yet
- White 1999Document9 pagesWhite 1999antonio damataNo ratings yet
- Hum. Mol. Genet. 1999 Raymond 989 96Document8 pagesHum. Mol. Genet. 1999 Raymond 989 96John MistryNo ratings yet
- Phylogenetic Structure of Q-M378 Subclade Based On Full Y-Chromosome SequencingDocument19 pagesPhylogenetic Structure of Q-M378 Subclade Based On Full Y-Chromosome SequencingVladimir GurianovNo ratings yet
- 1990 - Sequence-Specific DNA Binding by The C-Myc ProteinDocument3 pages1990 - Sequence-Specific DNA Binding by The C-Myc ProteinRaymond LaBoyNo ratings yet
- DNA Profiling Activity STUDENTDocument8 pagesDNA Profiling Activity STUDENTMonalisa SahuNo ratings yet
- Amblyrhynchus Cristatus 1Document4 pagesAmblyrhynchus Cristatus 1Arida FauziyahNo ratings yet
- Sim 2014Document2 pagesSim 2014Fausto PachecoNo ratings yet
- Genetic Linkage Map of The GuppyDocument14 pagesGenetic Linkage Map of The GuppyIsmael San Pablo GonzalezNo ratings yet
- 2016 Ruizruano Lmig Satellitome CompletoDocument42 pages2016 Ruizruano Lmig Satellitome CompletoDynhaNo ratings yet
- Band Methods 2Document7 pagesBand Methods 2Jazlynn WongNo ratings yet
- Mus Musculus Transformation Related Protein 73 (Trp73), Transcript Variant 3, mRNADocument7 pagesMus Musculus Transformation Related Protein 73 (Trp73), Transcript Variant 3, mRNAEeeeeeeeeNo ratings yet
- 3 WiseDocument10 pages3 WiseAnonymous 64x5K7Di9rNo ratings yet
- Morphological Variation in a Population of the Snake, Tantilla gracilis Baird and GirardFrom EverandMorphological Variation in a Population of the Snake, Tantilla gracilis Baird and GirardNo ratings yet
- (Plough: Drosophila Melanogaster'Document17 pages(Plough: Drosophila Melanogaster'Kaif Ali KhanNo ratings yet
- Genetic Distances Reconstruction of Phylogenetic Trees From Microsatellite DNADocument11 pagesGenetic Distances Reconstruction of Phylogenetic Trees From Microsatellite DNASabina TeofilovaNo ratings yet
- Karyotypic Studies - FullDocument8 pagesKaryotypic Studies - FullTJPRC PublicationsNo ratings yet
- Biology Shift 2 Nest 2023Document17 pagesBiology Shift 2 Nest 2023Hardik JoshiNo ratings yet
- The Trypanosoma Cruzi Genome Project: Nuclear Karyotype and Gene Mapping of Clone CL BrenerDocument8 pagesThe Trypanosoma Cruzi Genome Project: Nuclear Karyotype and Gene Mapping of Clone CL BrenerSergio Guillermo Cuevas CarbonellNo ratings yet
- Evolutionary Parameters of The Transcribed Mammalian GenomeDocument6 pagesEvolutionary Parameters of The Transcribed Mammalian GenomeLau Mandy KMNo ratings yet
- The Colugo (Cynocephalus Variegatus, Dermoptera) : The Primates' Gliding Sister?Document5 pagesThe Colugo (Cynocephalus Variegatus, Dermoptera) : The Primates' Gliding Sister?Samuel AlencastroNo ratings yet
- tmp475B TMPDocument18 pagestmp475B TMPFrontiersNo ratings yet
- Complete Genome Sequence of The Serotype K Streptococcus Mutans Strain LJ23Document2 pagesComplete Genome Sequence of The Serotype K Streptococcus Mutans Strain LJ23Nur Laila SariNo ratings yet
- New CellDocument60 pagesNew CellAna Paula CastroNo ratings yet
- Molecular Map of The Desmosomal PlaqueDocument12 pagesMolecular Map of The Desmosomal PlaqueLucas TobingNo ratings yet
- Zehner Et Al., 2004 - Genetic Identification of Forensically Important Flesh FliesDocument3 pagesZehner Et Al., 2004 - Genetic Identification of Forensically Important Flesh FliesJean-Bernard HuchetNo ratings yet
- CGR6Document28 pagesCGR6GenomicImage ProcessingNo ratings yet
- Assignment Biology (SB 015)Document6 pagesAssignment Biology (SB 015)Wei yangNo ratings yet
- Dna of Irish and Scottish SurnamesDocument37 pagesDna of Irish and Scottish Surnamesrgkelly62100% (2)
- Analiza Haplo Grupa Bosnjaka I Dr.Document12 pagesAnaliza Haplo Grupa Bosnjaka I Dr.damirzeNo ratings yet
- Significance of c1Q in Matriq BiologyDocument52 pagesSignificance of c1Q in Matriq BiologyKarthikeyini SokkalingamNo ratings yet
- Growth PredictionDocument64 pagesGrowth PredictionMothi KrishnaNo ratings yet
- Troubled WatersDocument150 pagesTroubled WatersABNo ratings yet
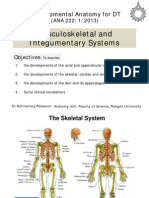
- Anatomy and Embryology of The Pharynx1.ppsDocument27 pagesAnatomy and Embryology of The Pharynx1.ppsНемосјановић ЋудмилаNo ratings yet
- LAB - The Axial Skeleton: ObjectivesDocument4 pagesLAB - The Axial Skeleton: ObjectivesSufiyaNo ratings yet
- JJJJJJJJJJJJJJJJJJ: FFFFFFFFFFFFFFFFFF FFFJJJFFJJFJFFJJFJDocument11 pagesJJJJJJJJJJJJJJJJJJ: FFFFFFFFFFFFFFFFFF FFFJJJFFJJFJFFJJFJAyansa ErgibaNo ratings yet
- Beastfolk RaceDocument2 pagesBeastfolk RaceAndrés MasNo ratings yet
- Upland Plover November-December 2018Document7 pagesUpland Plover November-December 2018NorthCountryBirdClubNo ratings yet
- Introductory Biology Worksheet1Document3 pagesIntroductory Biology Worksheet1Richie LewisNo ratings yet
- Systematics Based On Evolutionary Relationship 1Document13 pagesSystematics Based On Evolutionary Relationship 1Keith Penaso100% (3)
- Seçmeli Ingilizce Kısaltılmış2021Document4 pagesSeçmeli Ingilizce Kısaltılmış2021yaseminNo ratings yet
- แกะเทป Developmental of Musculoskeletal SystemDocument50 pagesแกะเทป Developmental of Musculoskeletal Systemjulesarojinee100% (1)
- Zoological Studies: An International JournalDocument142 pagesZoological Studies: An International JournalMizzannul HalimNo ratings yet
- Virtual Field Trip Report To Kopf Family Reservation July 2021Document41 pagesVirtual Field Trip Report To Kopf Family Reservation July 2021Western Cuyahoga AudubonNo ratings yet
- Zoo 211 (2018 - 2019) Past QuestionDocument3 pagesZoo 211 (2018 - 2019) Past Questionmusanafisat593No ratings yet
- Invading AntsDocument12 pagesInvading Antshamidreza mohseniNo ratings yet
- CarbohydratesDocument6 pagesCarbohydratesNarasimha MurthyNo ratings yet
- Tedersoo Et Al 2010 Ectomycorrhizal Lifestyle MycorrhizaDocument47 pagesTedersoo Et Al 2010 Ectomycorrhizal Lifestyle MycorrhizaFernando FabriniNo ratings yet
- Chapter 3 MIC260 PDFDocument36 pagesChapter 3 MIC260 PDFWan AiniNo ratings yet
- Modeling Limits To Cell SizeDocument6 pagesModeling Limits To Cell Sizeelorenzana0511No ratings yet
- Phylum Arthropoda: by DwiDocument40 pagesPhylum Arthropoda: by DwiNisa RNo ratings yet
- Brainy Klasa 4 DVD Worksheet U4Document1 pageBrainy Klasa 4 DVD Worksheet U4Gabriela KarpińskaNo ratings yet
- Plant MorphologyDocument34 pagesPlant Morphologyapi-262572717100% (2)
- General Zoology: Compilation of Laboratory ActivtesDocument17 pagesGeneral Zoology: Compilation of Laboratory ActivteshahahahaaaaaaaNo ratings yet
- Suci Dwi Wulandari (31) - XII MIPA 7 - Report Text UnderstandingDocument2 pagesSuci Dwi Wulandari (31) - XII MIPA 7 - Report Text UnderstandingSuci Dwi WulandariNo ratings yet
- Kindergarten Math and Critical Thinking WorksheetsDocument21 pagesKindergarten Math and Critical Thinking WorksheetsLisaNo ratings yet
- Chapter 6 - Cell Parts!: AP Biology Cells Review!!! Name - DateDocument2 pagesChapter 6 - Cell Parts!: AP Biology Cells Review!!! Name - DateMaryam AllanaNo ratings yet
- ĐÁNH VẦN TIẾNG ANH oxford phonics worldDocument30 pagesĐÁNH VẦN TIẾNG ANH oxford phonics worldoanh siuNo ratings yet
- Genetics Lesson PlanDocument12 pagesGenetics Lesson PlanYhan Brotamonte Boneo67% (9)
- Annotated Inventory of A Collection of Palauan Legends in The Belau National MuseumDocument14 pagesAnnotated Inventory of A Collection of Palauan Legends in The Belau National MuseumVanray TadaoNo ratings yet