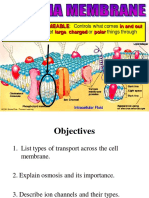
You might also like
- 2222 TransportDocument42 pages2222 TransportaddwrNo ratings yet
- 05 Membrane Transport VDocument27 pages05 Membrane Transport Vmaksud alaNo ratings yet
- Transport Across Membranes FinalDocument77 pagesTransport Across Membranes FinalLorraine Eve Abila100% (1)
- CH - 6, Passive TransportDocument43 pagesCH - 6, Passive TransportTony StarkNo ratings yet
- Physiology Seminar 1Document24 pagesPhysiology Seminar 1know more factsNo ratings yet
- Membrane Transport SystemsDocument33 pagesMembrane Transport SystemsObiajulu Crystal OnyinyechiNo ratings yet
- Lect 2 Membrane TransportDocument29 pagesLect 2 Membrane Transportramadan100% (1)
- Transport Function: of CellDocument41 pagesTransport Function: of CellDr. R. PeriasamyNo ratings yet
- Bio Grade 11 U-4 Note-2Document15 pagesBio Grade 11 U-4 Note-2betelhemwondimu421No ratings yet
- Transport Across Cell Membranes-1Document7 pagesTransport Across Cell Membranes-1Ifeoma JulietNo ratings yet
- Cell Membrane FunctionsDocument60 pagesCell Membrane FunctionsKate Clancy NugaoNo ratings yet
- 13-Movement of Substances Across The PDocument8 pages13-Movement of Substances Across The PAbbas TalibNo ratings yet
- Cytolog Membrtransport2 (Stud)Document4 pagesCytolog Membrtransport2 (Stud)OnSolomonNo ratings yet
- Transport Across The Cell Membrane - OdpDocument67 pagesTransport Across The Cell Membrane - OdpSilver CypressNo ratings yet
- Cell ExchangesDocument28 pagesCell ExchangesBenzerari ManaNo ratings yet
- Screenshot 2023-02-07 at 9.06.14 PMDocument92 pagesScreenshot 2023-02-07 at 9.06.14 PMMohammed RamzyNo ratings yet
- "In The Name Of,, .": AllahDocument28 pages"In The Name Of,, .": AllahAqsa ArifNo ratings yet
- Bio 122 Lec 1.2Document4 pagesBio 122 Lec 1.2Jara RogacionNo ratings yet
- Practical Work 3Document7 pagesPractical Work 3Achraf RabadiNo ratings yet
- Cell Lecture 5-1Document45 pagesCell Lecture 5-1gracechirangoNo ratings yet
- TransportDocument65 pagesTransportsiddhi meenaNo ratings yet
- Histology: Cytology, The Study of The Structure and Function of CellsDocument48 pagesHistology: Cytology, The Study of The Structure and Function of CellsZmeu CristinaNo ratings yet
- 1st Year 2nd Week PHYS Lecture 6 2019 2020Document14 pages1st Year 2nd Week PHYS Lecture 6 2019 2020Ahmed TarekNo ratings yet
- L6Document18 pagesL6AKNo ratings yet
- 2 TransportDocument36 pages2 Transportapi-19916399No ratings yet
- Chapter 2 - 1 - ForeignerDocument35 pagesChapter 2 - 1 - Foreignerapi-19916399No ratings yet
- BBT221 Lecture 2Document19 pagesBBT221 Lecture 2Al Sabri Bhuiyan 1812098042No ratings yet
- Chapter 3: Movement of Substances Across The Plasma MembraneDocument28 pagesChapter 3: Movement of Substances Across The Plasma MembraneMohd Asraf90% (10)
- 4 - Membrane PhysiologyDocument31 pages4 - Membrane PhysiologysadaffardoosNo ratings yet
- Membrane Transport ABDocument38 pagesMembrane Transport ABdj77djcqy7No ratings yet
- Physiology L2Document13 pagesPhysiology L2dfffggg2je443qfdfvhkhNo ratings yet
- Movements Across Cell MembraneDocument30 pagesMovements Across Cell Membraneعلیزہ نعیمNo ratings yet
- MEMBRANE TRANSPORTand DISEASES MBBSDocument48 pagesMEMBRANE TRANSPORTand DISEASES MBBSAbdulateef AdebisiNo ratings yet
- Membrane Tranport 2ND Lecture by Dr. RoomiDocument32 pagesMembrane Tranport 2ND Lecture by Dr. RoomiMudassar Roomi100% (2)
- Topic 1.4 Membrane TransportDocument8 pagesTopic 1.4 Membrane Transportjasmine wibawaNo ratings yet
- Lecture On Membrane Transport by Dr. Mudassar Ali Roomi.Document37 pagesLecture On Membrane Transport by Dr. Mudassar Ali Roomi.Mudassar Roomi100% (1)
- Cell TransportDocument18 pagesCell TransportCallista FeliciaNo ratings yet
- Active Transport MethodsDocument13 pagesActive Transport Methodssrikanth100% (3)
- Biochem Lab For ReadingDocument57 pagesBiochem Lab For ReadingShaira Elyze GabrielNo ratings yet
- Biophysics-Lectures-2012-3 (Physiology) PDFDocument107 pagesBiophysics-Lectures-2012-3 (Physiology) PDFNedelcuGeorgeNo ratings yet
- Physiology U-2 Transport Across The Plasma MembraneDocument48 pagesPhysiology U-2 Transport Across The Plasma Membranesinte beyuNo ratings yet
- Difusi OsmosisDocument53 pagesDifusi OsmosisSulis tyani08No ratings yet
- Transport Across Cell MembraneDocument26 pagesTransport Across Cell MembraneAlhadaeel JesjNo ratings yet
- 1-Passive Transport:: Selectively Permeable BarriersDocument19 pages1-Passive Transport:: Selectively Permeable Barriersapi-3706483No ratings yet
- Nerve and Muscle PhysiologyDocument82 pagesNerve and Muscle PhysiologychandsriNo ratings yet
- Transport Across MembranesDocument61 pagesTransport Across MembranesAmara TargaryenNo ratings yet
- Transport 2 PDF Lecture Elalamein 2021-22Document5 pagesTransport 2 PDF Lecture Elalamein 2021-22Ahmed MagdyNo ratings yet
- Physio Movement by Cell MembraneDocument30 pagesPhysio Movement by Cell MembranemnaeditzNo ratings yet
- Membrane Physiology: Human Physiology By-Liul M. ForDocument34 pagesMembrane Physiology: Human Physiology By-Liul M. ForKhant Si ThuNo ratings yet
- Transport Cell: Diana N. AffandiDocument36 pagesTransport Cell: Diana N. AffandiunyeNo ratings yet
- 1.0 The Origin of Biopotentials (New)Document62 pages1.0 The Origin of Biopotentials (New)Hanif HussinNo ratings yet
- 2 CellsDocument33 pages2 CellsAn ThịnhNo ratings yet
- Cell Physiology 1.Document3 pagesCell Physiology 1.Raj SinghNo ratings yet
- CH5 Membrane TraffickingDocument48 pagesCH5 Membrane TraffickingAhmed MohamadNo ratings yet
- Cellular Transport ConceptDocument7 pagesCellular Transport ConceptRuhul Qudus NaimNo ratings yet
- Raven Biology of Plants: Eighth EditionDocument29 pagesRaven Biology of Plants: Eighth EditionMoath EnnabNo ratings yet
- 1 4Document10 pages1 4nevio.wirtgenNo ratings yet
- CellularTransport MATSAPUMEDocument37 pagesCellularTransport MATSAPUMEPanyapat SrisawatNo ratings yet
- ACTIVE TRANSPORT Vs PASSIVE TRANSPORTDocument6 pagesACTIVE TRANSPORT Vs PASSIVE TRANSPORTfelixnellyNo ratings yet
- Biology: a QuickStudy Laminated Reference GuideFrom EverandBiology: a QuickStudy Laminated Reference GuideRating: 3 out of 5 stars3/5 (2)
- Science of LIVING SYSTEM AB (Part of Unit 2)Document46 pagesScience of LIVING SYSTEM AB (Part of Unit 2)himanshu_agraNo ratings yet
- R-Sen Lecture 01aDocument28 pagesR-Sen Lecture 01ahimanshu_agraNo ratings yet
- RF Module Quick Mannua v3Document3 pagesRF Module Quick Mannua v3himanshu_agraNo ratings yet
- BS 3Document60 pagesBS 3himanshu_agraNo ratings yet
- Motifs of Protein StructureDocument41 pagesMotifs of Protein Structurehimanshu_agraNo ratings yet
- Science of Living Systems: Bio-Thermal-Fluid Sciences: Lecture by Prof. Soumen DasDocument36 pagesScience of Living Systems: Bio-Thermal-Fluid Sciences: Lecture by Prof. Soumen Dashimanshu_agraNo ratings yet
- Science of Living SystemsDocument52 pagesScience of Living Systemshimanshu_agraNo ratings yet
- Bioscience: Protein Structure Function BS20001 Section 2Document27 pagesBioscience: Protein Structure Function BS20001 Section 2himanshu_agraNo ratings yet
- Waste Minimization and Cleaner ProductionDocument75 pagesWaste Minimization and Cleaner Productionhimanshu_agra100% (2)
- Soild Waste AmangementDocument75 pagesSoild Waste Amangementhimanshu_agraNo ratings yet
- Photosynthesis: Prof. S. Dutta GuptaDocument41 pagesPhotosynthesis: Prof. S. Dutta Guptahimanshu_agraNo ratings yet
- Noise PolutionDocument48 pagesNoise Polutionhimanshu_agraNo ratings yet
- Evs - Soil PollutionDocument47 pagesEvs - Soil Pollutionhimanshu_agraNo ratings yet
- Chapter 7: Entity-Relationship Model: Database System Concepts, 6 EdDocument102 pagesChapter 7: Entity-Relationship Model: Database System Concepts, 6 Edhimanshu_agraNo ratings yet
- Wastewater Treatment: Department of Civil Engineering Indian Institute of Technology KharagpurDocument26 pagesWastewater Treatment: Department of Civil Engineering Indian Institute of Technology Kharagpurhimanshu_agraNo ratings yet
- Dart Language SpecificationDocument111 pagesDart Language SpecificationLiaboNo ratings yet
- EliminationDocument12 pagesEliminationMatthew JohnNo ratings yet
- Year 2 WeeblyDocument1 pageYear 2 Weeblyapi-257776108No ratings yet
- Tugas Paper Mekanika PatahanDocument1 pageTugas Paper Mekanika PatahanRahmandhika Firdauzha Hary HernandhaNo ratings yet
- How To Draw Super-Powered Heroes Vol. 1Document129 pagesHow To Draw Super-Powered Heroes Vol. 1LuisFernandoRocaMejia88% (33)
- Manuel Guindeau X2Document53 pagesManuel Guindeau X2fr_emailleNo ratings yet
- Chapter 4 Semantic Roles - NOTESDocument8 pagesChapter 4 Semantic Roles - NOTESWaad MajidNo ratings yet
- Chains For Power TransmissionDocument11 pagesChains For Power TransmissionQasim JavedNo ratings yet
- P 278 - GN 1007Document25 pagesP 278 - GN 1007Doben DalisayNo ratings yet
- AH Saphiro 3 RD Chapter PDFDocument26 pagesAH Saphiro 3 RD Chapter PDFRahul RoyNo ratings yet
- Of Intrinsic Validity - A Study On The Relevance of Pūrva Mīmā SāDocument29 pagesOf Intrinsic Validity - A Study On The Relevance of Pūrva Mīmā SāDanillo Costa LimaNo ratings yet
- Respiratory System: StructureDocument29 pagesRespiratory System: StructureDr. Abir Ishtiaq100% (1)
- Chemistry Unit Lesson Plans Week 13Document2 pagesChemistry Unit Lesson Plans Week 13Janix MagbanuaNo ratings yet
- Chapter 09 - FracturePressureDocument55 pagesChapter 09 - FracturePressuredwiokkysaputraNo ratings yet
- Product Manual: Panel Mounted ControllerDocument271 pagesProduct Manual: Panel Mounted ControllerLEONARDO FREITAS COSTANo ratings yet
- Petrel 2013 Property Modeling Course: Module 2: Data PreparationDocument15 pagesPetrel 2013 Property Modeling Course: Module 2: Data PreparationJaninne Campo100% (1)
- IMO 2019 NotesDocument13 pagesIMO 2019 NotesAndrei346No ratings yet
- Euler-Maclaurin Expansion and Woolhouse's Formula: A S. M Volume 2, Pp. 631-633 inDocument3 pagesEuler-Maclaurin Expansion and Woolhouse's Formula: A S. M Volume 2, Pp. 631-633 inAyu Nurul AtiqohNo ratings yet
- Maths Formula's Pocket BookDocument113 pagesMaths Formula's Pocket BookPOLUNATI SAHITHI POLUNATI SAHITHI100% (1)
- Solutions - JEE Main 2024 January Question Bank - MathonGoDocument7 pagesSolutions - JEE Main 2024 January Question Bank - MathonGoLeo UzumakiNo ratings yet
- A Syntactic Analysis of Surah Al MasadDocument14 pagesA Syntactic Analysis of Surah Al MasadLisda Farikhatunnisak100% (1)
- QUCM CCM Application ManualDocument30 pagesQUCM CCM Application ManualibanvegaNo ratings yet
- Jntu Citd M.tech Mechatronics SyllabusDocument24 pagesJntu Citd M.tech Mechatronics SyllabusSai KiranNo ratings yet
- Rectangular Flange DesignDocument6 pagesRectangular Flange DesignTAHIR MAHMOODNo ratings yet
- Project 4Document11 pagesProject 4Kara WarnerNo ratings yet
- IWEX - A New Ultrasonic Array Technology For Direct Imaging of Subsurface DefectsDocument5 pagesIWEX - A New Ultrasonic Array Technology For Direct Imaging of Subsurface DefectsIndigo WaveNo ratings yet
- MaxwellDocument2 pagesMaxwellMAURICENo ratings yet
- Explosive Welding 1 WeldingDocument4 pagesExplosive Welding 1 WeldingRabindra DashNo ratings yet
- Module 6 - Introduction To Hypertext PreprocessorDocument19 pagesModule 6 - Introduction To Hypertext PreprocessorMr. EpiphanyNo ratings yet
- Lab Exe No. 10 - Design of Simple Bioethanol Production System - FermentationDocument2 pagesLab Exe No. 10 - Design of Simple Bioethanol Production System - FermentationJamil Mansor M. LaguiabNo ratings yet