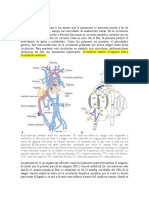
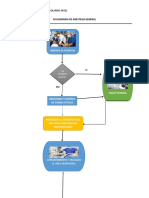
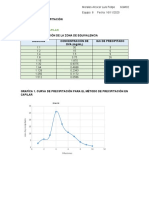
You might also like
- Metabolismo Del ArnDocument9 pagesMetabolismo Del ArnNathalie CortezNo ratings yet
- PDF Triptico Tamiz NeonatalDocument2 pagesPDF Triptico Tamiz NeonatalMeliza MeraNo ratings yet
- Investigación y Confirmación de Brotes-2Document17 pagesInvestigación y Confirmación de Brotes-2Leticia ConstantinoNo ratings yet
- Cap. 8 - Psicología y Promoción Salud El Caso de La Alimentación InfantilDocument82 pagesCap. 8 - Psicología y Promoción Salud El Caso de La Alimentación InfantilAnto CabonaNo ratings yet
- Fisiologia FetalDocument13 pagesFisiologia Fetalmanibea100% (1)
- Aminas SimpaticomimeticasDocument13 pagesAminas SimpaticomimeticasCarolina F' RochaNo ratings yet
- Las Paperas o Parotiditis Epidemica PDFDocument7 pagesLas Paperas o Parotiditis Epidemica PDFmanuelNo ratings yet
- Hdne Giardia LambliaDocument2 pagesHdne Giardia LambliaCarlos ChavezNo ratings yet
- Norma 197Document7 pagesNorma 197Karla Andrea JonasNo ratings yet
- 3er Tema Enfermedades Agudas Transmisibles y No Transmisibles.Document25 pages3er Tema Enfermedades Agudas Transmisibles y No Transmisibles.Daiane BorgoNo ratings yet
- Intolerancia A La Lactosa y Azúcares ReductoresDocument21 pagesIntolerancia A La Lactosa y Azúcares ReductoresMarianaRincon0% (1)
- Los Encimas Como Sitios de Acción de Los FármacosDocument5 pagesLos Encimas Como Sitios de Acción de Los FármacosDOminique Venemeti100% (1)
- La Tríada Ecológica ?Document8 pagesLa Tríada Ecológica ?Karen MacuareNo ratings yet
- Historia Natural de La Toxoplasmosis 1Document20 pagesHistoria Natural de La Toxoplasmosis 1AngelNZNo ratings yet
- Circulación Fetal - PediatriaDocument5 pagesCirculación Fetal - Pediatriaomar duvan vargas moralesNo ratings yet
- Sondas VesicalesDocument17 pagesSondas VesicalesAbdelaziz El Aakrout Kaamouchi100% (1)
- Triptico-Necator AmericanusDocument2 pagesTriptico-Necator AmericanusANdriid Lopez ClavijoNo ratings yet
- PulicosisDocument3 pagesPulicosisjackriden100% (1)
- Fagocitosis (Microbiología)Document3 pagesFagocitosis (Microbiología)Andrea Huerta EscobedoNo ratings yet
- ÓRGANO DIANA - Qué Es, Definición y EjemplosDocument7 pagesÓRGANO DIANA - Qué Es, Definición y EjemplosEnrique AlvarezNo ratings yet
- ARO GinecoDocument39 pagesARO GinecoHellen GordilloNo ratings yet
- Ciclo Vital de La Familia y El IndividuoDocument24 pagesCiclo Vital de La Familia y El IndividuoRommicitaNo ratings yet
- Clasificación CatéteresDocument7 pagesClasificación CatéteresDeysisita Ruiz CamposNo ratings yet
- Caso Clínico 2Document5 pagesCaso Clínico 2ErickAlvarezNo ratings yet
- Valoracion de La EmbarazadaDocument14 pagesValoracion de La Embarazadaerika2609No ratings yet
- Signos Vitales POR GRUPO ETAREODocument43 pagesSignos Vitales POR GRUPO ETAREOROSSY VARGASNo ratings yet
- Atencion Inmediata Al Recien NacidoDocument18 pagesAtencion Inmediata Al Recien NacidoEnilce Jessica CruzNo ratings yet
- AnencefaliaDocument13 pagesAnencefaliaGISELA ORTIZNo ratings yet
- Manual de VacunaciónDocument29 pagesManual de VacunaciónVillarreal Galán Andrea Jazmin100% (1)
- PlaceDocument3 pagesPlacejudith velzNo ratings yet
- Desinfección de La Sala QuirúrgicaDocument18 pagesDesinfección de La Sala QuirúrgicaCristhian De Jesus Mendez HernandezNo ratings yet
- Diágrama de FlujoDocument3 pagesDiágrama de FlujoKaren Briseño CabreraNo ratings yet
- Place EclampsiaDocument2 pagesPlace EclampsiaVioleta AlmarazNo ratings yet
- Historia Natural y Social de La ParasitosisDocument5 pagesHistoria Natural y Social de La Parasitosisrene clausi salazar100% (1)
- FAMILIOGRAMADocument32 pagesFAMILIOGRAMAAlfonsoSaldañaChuquizutaNo ratings yet
- De Acuerdo Con La OMSDocument7 pagesDe Acuerdo Con La OMSHeberVelasquezNo ratings yet
- Nom 003 Ssa2 1993Document20 pagesNom 003 Ssa2 1993César Serrano MaciasNo ratings yet
- Historia Natural Embarazo EctópicoDocument3 pagesHistoria Natural Embarazo EctópicoYolanda Paola MuvdiNo ratings yet
- Selección de Datos para La VigilanciaDocument22 pagesSelección de Datos para La VigilanciaKathyMartínezNo ratings yet
- Seminario Metabolismo ADN 2016Document51 pagesSeminario Metabolismo ADN 2016Melanie TassoNo ratings yet
- Llinea Del Tiempo Cuidado Paliativo - OrganizedDocument3 pagesLlinea Del Tiempo Cuidado Paliativo - OrganizedVanesa FloresNo ratings yet
- Mapa ConceptualDocument2 pagesMapa Conceptualapi-472592451No ratings yet
- Museo de Medicina EnsayoDocument4 pagesMuseo de Medicina EnsayoMichelle MarinNo ratings yet
- Unam Practica Gasometria y DialisisDocument14 pagesUnam Practica Gasometria y DialisisJesus CervantesNo ratings yet
- Historia Clinica FORMATO BlancoDocument6 pagesHistoria Clinica FORMATO BlancoNayro Gaym Gaza CortesNo ratings yet
- Tira HorariaDocument15 pagesTira HorariaTriny ValdezNo ratings yet
- Control TermicoDocument22 pagesControl TermicoGabriela Castillo González100% (1)
- Norma Oficial Mexicana NOM-007 - SSA2-2016, para La Atención de La Mujer Durante El Embarazo, Parto y Puerperio, y de La Persona Recién Nacida.Document28 pagesNorma Oficial Mexicana NOM-007 - SSA2-2016, para La Atención de La Mujer Durante El Embarazo, Parto y Puerperio, y de La Persona Recién Nacida.IRMA ADELAIDA ACOSTA CASTRONo ratings yet
- Detección Temprana y Atención Oportuna de Enfermedades CongenitasDocument15 pagesDetección Temprana y Atención Oportuna de Enfermedades CongenitasRendón Magali100% (1)
- Neumococo 13 y 23Document2 pagesNeumococo 13 y 23Carlos VazquezNo ratings yet
- FLUJOGRAMA DE ANESTESIA GENERAL Univ GUZMAN FLORES ROLANDO JHOEL 3DDocument4 pagesFLUJOGRAMA DE ANESTESIA GENERAL Univ GUZMAN FLORES ROLANDO JHOEL 3DRolando Jhoel Guzman100% (1)
- Depresión PlaceDocument11 pagesDepresión Placekaren jennifer chavez jimenezNo ratings yet
- Cronograma de Actividades EnfemeriaDocument2 pagesCronograma de Actividades Enfemeriaeric jesusNo ratings yet
- Epidemiologia de Herpes GenitalDocument4 pagesEpidemiologia de Herpes GenitalIngrid CanizNo ratings yet
- 2 Cuidados Del Neonato en IncubadoraDocument21 pages2 Cuidados Del Neonato en IncubadoraLuis Palacios0% (1)
- Historia Natural de La ApendicitisDocument8 pagesHistoria Natural de La ApendicitisMaríaFernandaVelascoNo ratings yet
- Cuadro Comparativo MicrobiologiaDocument3 pagesCuadro Comparativo MicrobiologiaMonica PerdomoNo ratings yet
- Introduccion A La PediatriaDocument22 pagesIntroduccion A La PediatriaDaiiana CentenoNo ratings yet
- Proceso de TranscripciónDocument17 pagesProceso de TranscripciónKarina Alejandra Cervantes RoblesNo ratings yet
- ARN y Sus CaracterísticasDocument41 pagesARN y Sus CaracterísticasNazareth EspinozaNo ratings yet
- Cuestionario 05 - Bioca PrácticaDocument5 pagesCuestionario 05 - Bioca PrácticaMarjorie Lizbeth P.No ratings yet
- Taller de Replicacion y TraduccionDocument4 pagesTaller de Replicacion y TraduccionNataliaCharryNo ratings yet
- Resumenes de BBM 2021 Por Denise BourlotDocument270 pagesResumenes de BBM 2021 Por Denise BourlotEmylyn AndradeNo ratings yet
- Las BiomoléculasDocument25 pagesLas BiomoléculasDe Val RickNo ratings yet
- Secuenciación de ProteínasDocument13 pagesSecuenciación de ProteínasManuel A. VelazNo ratings yet
- Resumen Capitulo 4 LehningerDocument4 pagesResumen Capitulo 4 LehningerÉrick JuárezNo ratings yet
- Nucleotidos y NucleoprotreinasDocument38 pagesNucleotidos y NucleoprotreinasNazareth LucianaNo ratings yet
- Informe de Laboratorio Actividad EnzimaticaDocument6 pagesInforme de Laboratorio Actividad EnzimaticaLUIS ANGEL LLANOS ACUÑANo ratings yet
- 20 Tipos de Proteínas y Sus Funciones en El CuerpoDocument1 page20 Tipos de Proteínas y Sus Funciones en El CuerpoGuereca Montelongo Andrea100% (1)
- Examen de BioquimicaDocument2 pagesExamen de BioquimicaYuMi BautistaNo ratings yet
- Factores Físicos y Químicos Que Alteran La Función y Estructura de Las ProteínasDocument3 pagesFactores Físicos y Químicos Que Alteran La Función y Estructura de Las ProteínasCristian Contreras100% (2)
- Bioquimica Pensum IVICDocument1 pageBioquimica Pensum IVICMiguel León RoperoNo ratings yet
- Diagrama de LanzaderasDocument1 pageDiagrama de LanzaderasedipsonNo ratings yet
- Taller Sintesis de Proteinas 2020Document3 pagesTaller Sintesis de Proteinas 2020Luis Blanco Blanco gomez100% (1)
- Metabolismo de Aminoacidos y ProteinasDocument49 pagesMetabolismo de Aminoacidos y Proteinasjuan100% (1)
- 15 Preguntas Acerca de Las ProteínasDocument4 pages15 Preguntas Acerca de Las ProteínasAndresfdezcmc94% (47)
- Reaccines de PrecipitaciónDocument4 pagesReaccines de PrecipitaciónLuis AlcocerNo ratings yet
- Test Diagnóstico de BioquímicaDocument5 pagesTest Diagnóstico de BioquímicaAbril Tames FerrufinoNo ratings yet
- Genética UCSUR - T1 - Bases Moleculares de La HerenciaDocument57 pagesGenética UCSUR - T1 - Bases Moleculares de La HerenciaDiego Cruzado oroscoNo ratings yet
- ADN, Genes y Código GenéticoDocument6 pagesADN, Genes y Código GenéticomoreNo ratings yet
- CASEÍNADocument9 pagesCASEÍNAmjambrinaNo ratings yet
- Programa Diseño de BioproductosDocument3 pagesPrograma Diseño de BioproductosLauraNo ratings yet
- Biosíntesisde Ácidos GrasosDocument88 pagesBiosíntesisde Ácidos GrasosUSMP FN ARCHIVOSNo ratings yet
- Preguntas ProteinasDocument2 pagesPreguntas ProteinasandreaNo ratings yet
- Guia 5 TAP BiologíaDocument9 pagesGuia 5 TAP Biología7ndym5dj8vNo ratings yet
- Química BiológicaDocument35 pagesQuímica BiológicaNeira Osorio100% (1)
- Proteosomas y UbiquitinaDocument23 pagesProteosomas y UbiquitinaGabriela Rodriguez MuñozNo ratings yet
- Biomoleculas David Velasco Tarea 3Document6 pagesBiomoleculas David Velasco Tarea 3David GoyesNo ratings yet
- Anual SM Semana 03 - BiologíaDocument19 pagesAnual SM Semana 03 - BiologíaGeordan SanchezNo ratings yet
- Membrana PlasmáticaDocument5 pagesMembrana PlasmáticaMazuey CroissantsNo ratings yet