You might also like
- GRUPO 5 Bioenergetica-1Document29 pagesGRUPO 5 Bioenergetica-1Nicole Garavito NuñezNo ratings yet
- Bioquímica: Enzimas Oxido-Reductasas Involucradas en Reacciones RedoxDocument24 pagesBioquímica: Enzimas Oxido-Reductasas Involucradas en Reacciones RedoxAnonimo22No ratings yet
- Resumen Capitulos Bioquimi1ca PDFDocument36 pagesResumen Capitulos Bioquimi1ca PDFRuben ReynosoNo ratings yet
- Cadena RespiratoriaDocument60 pagesCadena RespiratoriaYoselin RomeroNo ratings yet
- Reacciones MetabólicasDocument14 pagesReacciones MetabólicasJoel PolakoNo ratings yet
- Clase 3 - Bioquimica - DR Zarate 2023 Oxidaciones Biolog.Document22 pagesClase 3 - Bioquimica - DR Zarate 2023 Oxidaciones Biolog.Angie Alvarez BurgosNo ratings yet
- Reacciones Oxido Reduccion BioquimicaDocument33 pagesReacciones Oxido Reduccion BioquimicaLaura LopezNo ratings yet
- Resumen Solemne 2 BiologiaDocument9 pagesResumen Solemne 2 BiologiaaylinesotorNo ratings yet
- Partes TeoríaDocument3 pagesPartes TeoríaTimana Cruz Jesús JoséNo ratings yet
- BIOENERGETICADocument8 pagesBIOENERGETICALucimar SosaNo ratings yet
- 2 Oxidaciones BiologicasDocument9 pages2 Oxidaciones BiologicasLucasNo ratings yet
- Complejos de Cadena ReespDocument9 pagesComplejos de Cadena ReespMicaela AliNo ratings yet
- Inhibidores y Desacopladores de La Cadena Respiratoria... 3º Ciclo ... AgronomíaDocument5 pagesInhibidores y Desacopladores de La Cadena Respiratoria... 3º Ciclo ... AgronomíaDarli Alvaro Camacho RojasNo ratings yet
- Monografia - Cadena Respiratoria y Fosforilación OxidativaDocument4 pagesMonografia - Cadena Respiratoria y Fosforilación OxidativaNoemi RadaNo ratings yet
- MetabolismoDocument7 pagesMetabolismoFatima Serrano PerezNo ratings yet
- Metabolismo CatabolismoDocument33 pagesMetabolismo CatabolismoFrancis A. Batarse0% (1)
- Informe Temario Bioquímica Ii 2021-2022 Primer ParcialDocument13 pagesInforme Temario Bioquímica Ii 2021-2022 Primer ParcialCinthya BenavidesNo ratings yet
- Respiracion CelularDocument82 pagesRespiracion CelularDaylibeth CamperoNo ratings yet
- 10 Energía 2016Document8 pages10 Energía 2016Marcelo EscalanteNo ratings yet
- Bioquimica II, Bioenergetica y MetabolismoDocument40 pagesBioquimica II, Bioenergetica y MetabolismoOneida MorenoNo ratings yet
- Metabolismo celular: glucólisis y ciclo de KrebsDocument61 pagesMetabolismo celular: glucólisis y ciclo de KrebsAndres Monroy100% (1)
- Presentacion 1 Bioenergetica 13 de MayoDocument23 pagesPresentacion 1 Bioenergetica 13 de MayoJubitza Paredes MamaniNo ratings yet
- Respiración Celular - GlosarioDocument3 pagesRespiración Celular - GlosarioCarlos ChiaNo ratings yet
- Fosforilacion Oxidativa Teoria QuimiosmoticaDocument18 pagesFosforilacion Oxidativa Teoria QuimiosmoticaJose Arceo CendejasNo ratings yet
- Cadena RespiratoriaDocument42 pagesCadena RespiratoriaMacedo Garcia Henry EmptyNo ratings yet
- Ganancia Total Del ATP en La Respiración Celular AeróbicaDocument6 pagesGanancia Total Del ATP en La Respiración Celular AeróbicaNataly MonterosNo ratings yet
- Cadena Respiratoria Aporte Elctronico y Fosforilación OxidativaDocument40 pagesCadena Respiratoria Aporte Elctronico y Fosforilación Oxidativa悲しい エレン・イェーガーNo ratings yet
- Unidad Ii. Cadena Respiratoria y Fosforilacion Oxidativa PDFDocument34 pagesUnidad Ii. Cadena Respiratoria y Fosforilacion Oxidativa PDFMafer AraujoNo ratings yet
- Metabolismo Gluco Kbres MedicinaDocument40 pagesMetabolismo Gluco Kbres MedicinaLuis eduardo La torre AdrianzénNo ratings yet
- Principales Rutas CatabólicasDocument49 pagesPrincipales Rutas CatabólicasReben ZecnajNo ratings yet
- Oxidación biológica: flujo de electronesDocument16 pagesOxidación biológica: flujo de electronesDelmira Andia challcoNo ratings yet
- Práctica N4 BioquimicaDocument14 pagesPráctica N4 BioquimicaJose FernandoNo ratings yet
- OXIDACIÓN de NUTRIENTES y PRODUCCIÓN de ENERGIADocument3 pagesOXIDACIÓN de NUTRIENTES y PRODUCCIÓN de ENERGIAsilrox3111No ratings yet
- Resumen BioquimicaDocument18 pagesResumen BioquimicaMary Elizabeth Bravo Peña100% (2)
- BioquimicaDocument3 pagesBioquimicaP. Arevalo IlianaNo ratings yet
- Seminario 1Document4 pagesSeminario 1Renzzo Amaro Mozo MarquinaNo ratings yet
- Metabolismo CelularDocument45 pagesMetabolismo CelularMilagros MuñizNo ratings yet
- Transferencia de energía en la bioenergética celularDocument21 pagesTransferencia de energía en la bioenergética celularsara manriquezNo ratings yet
- Carbohidratos y lípidos: bioenergética y metabolismoDocument55 pagesCarbohidratos y lípidos: bioenergética y metabolismoSusana CNo ratings yet
- Respiración AeróbicaDocument33 pagesRespiración AeróbicaCARLOS VERGARA COQUENo ratings yet
- Procesos metabólicos celulares: glucólisis, ciclo de Krebs, cadena respiratoria y ATP sintasaDocument32 pagesProcesos metabólicos celulares: glucólisis, ciclo de Krebs, cadena respiratoria y ATP sintasaFan Do.No ratings yet
- Respiracion en PLantasDocument82 pagesRespiracion en PLantasJesús LunarNo ratings yet
- Semana 4 - Metabolismo y Produccion de EnergiaDocument9 pagesSemana 4 - Metabolismo y Produccion de EnergianicolasmacarisNo ratings yet
- Respiración vegetal y resistencia al cianuroDocument6 pagesRespiración vegetal y resistencia al cianuroJhenner Ivan Mendoza PerezNo ratings yet
- Metabolism oDocument16 pagesMetabolism oMia MontgomeryNo ratings yet
- Oxidacion BiologicaDocument25 pagesOxidacion BiologicaBeberly Reyes EspinozaNo ratings yet
- BIOQUIMICA CLASE 2: 14 de OctubreDocument21 pagesBIOQUIMICA CLASE 2: 14 de OctubreanaNo ratings yet
- Sintesis de CarbohidratosDocument53 pagesSintesis de CarbohidratosJR LindoNo ratings yet
- Metabolismo BacterianoDocument46 pagesMetabolismo BacterianoAndres Flores PardaveNo ratings yet
- Glosario BioquimicoDocument12 pagesGlosario BioquimicoAngelitho Muñoz ChavezNo ratings yet
- Metabolismo energético: Reacciones catabólicas y anabólicasDocument40 pagesMetabolismo energético: Reacciones catabólicas y anabólicasCristhian Nc100% (2)
- Respiracion Celular Unidad 10Document7 pagesRespiracion Celular Unidad 10valentinaNo ratings yet
- Una Forma Más General de Interpretar Las Reacciones ÓxidoDocument3 pagesUna Forma Más General de Interpretar Las Reacciones ÓxidoAlex Hdez AlfaroNo ratings yet
- Bioquímica 15 Tarea3Document13 pagesBioquímica 15 Tarea3juliana22004No ratings yet
- CadenaRespiratoria7EnzimasDocument22 pagesCadenaRespiratoria7EnzimasStiven Roldan ParedesNo ratings yet
- Folleto de BioquimicaDocument496 pagesFolleto de BioquimicaHildebrando Pablo Ataco CarrionNo ratings yet
- Acceso a Universidad para Mayores de 25 años. Biología 2013-2017.: Solucionario Pruebas 2013-2017From EverandAcceso a Universidad para Mayores de 25 años. Biología 2013-2017.: Solucionario Pruebas 2013-2017No ratings yet
- Acceso a Universidad para Mayores de 25 años. Biología.: Solucionario Pruebas 2006-2012From EverandAcceso a Universidad para Mayores de 25 años. Biología.: Solucionario Pruebas 2006-2012No ratings yet
- Volumenes de Produccion de VinoDocument4 pagesVolumenes de Produccion de VinoMaricarmenNo ratings yet
- CerealesDocument94 pagesCerealesMaricarmenNo ratings yet
- Análisis sensorial de la miel de abejaDocument8 pagesAnálisis sensorial de la miel de abejaMaricarmenNo ratings yet
- Trazas MineralesDocument19 pagesTrazas MineralesMaricarmenNo ratings yet
- Analisis de GrasasDocument4 pagesAnalisis de GrasasMaricarmenNo ratings yet
- CerealesDocument94 pagesCerealesMaricarmenNo ratings yet
- Determinacón de ProteínasDocument5 pagesDeterminacón de ProteínasMaricarmenNo ratings yet
- Residuos Solidos 2017Document71 pagesResiduos Solidos 2017MaricarmenNo ratings yet
- Contaminacion Del Agua-1Document38 pagesContaminacion Del Agua-1MaricarmenNo ratings yet
- Resumen en PDF Evaporacion PDFDocument38 pagesResumen en PDF Evaporacion PDFMaricarmenNo ratings yet
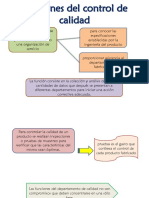
- Funciones Del Control de CalidadDocument5 pagesFunciones Del Control de CalidadMaricarmenNo ratings yet
- Tratamiento de Aguas NaturalesDocument16 pagesTratamiento de Aguas NaturalesMaricarmenNo ratings yet
- Localizacion de Valor FuturoDocument8 pagesLocalizacion de Valor FuturoMaricarmenNo ratings yet
- Contaminacion Del SueloDocument26 pagesContaminacion Del SueloMaricarmenNo ratings yet
- Análisis físico de aceites y grasasDocument5 pagesAnálisis físico de aceites y grasasMaricarmenNo ratings yet
- Higiene y Saneamiento en La Industria AlimentariaDocument36 pagesHigiene y Saneamiento en La Industria AlimentariaMaricarmenNo ratings yet
- Control AmbientalDocument6 pagesControl AmbientalMaricarmenNo ratings yet
- Maquinaria básica bodega vinoDocument17 pagesMaquinaria básica bodega vinoMaricarmen0% (1)
- CarneDocument30 pagesCarneMaricarmenNo ratings yet
- Diapos de MuestreoDocument28 pagesDiapos de MuestreoMaricarmenNo ratings yet
- Carnicos PatéDocument28 pagesCarnicos PatéMaricarmenNo ratings yet
- Tema 2Document16 pagesTema 2MaricarmenNo ratings yet
- Vizcacha-Parte - 3 y 5Document7 pagesVizcacha-Parte - 3 y 5MaricarmenNo ratings yet
- EnologiaDocument51 pagesEnologiaMaricarmenNo ratings yet
- Diapos Originales de AnalisisDocument28 pagesDiapos Originales de AnalisisMaricarmenNo ratings yet
- Agua LibreDocument2 pagesAgua LibreMaricarmenNo ratings yet
- Ing. Héctor Lorenzo Villa CajavilcaDocument36 pagesIng. Héctor Lorenzo Villa CajavilcaRicardoNo ratings yet
- DiaposDocument5 pagesDiaposMaricarmenNo ratings yet
- Recuperación y Purificación de Productos BilógicosDocument10 pagesRecuperación y Purificación de Productos BilógicosMaricarmenNo ratings yet
- Costos de Calidad y No Calidad y Mejoramiento de ProcesosDocument12 pagesCostos de Calidad y No Calidad y Mejoramiento de ProcesosMaricarmen50% (2)
- Urtica Dioica - Wikipedia, La Enciclopedia LibreDocument13 pagesUrtica Dioica - Wikipedia, La Enciclopedia LibreCésar RiveraNo ratings yet
- El Misterio de La Bruja Aullante - OneShot Del Sot Equus (Final)Document14 pagesEl Misterio de La Bruja Aullante - OneShot Del Sot Equus (Final)Joan P. PereiraNo ratings yet
- El Tratamiento de Los Medios Probatorios en La Nueva Ley Procesal Del TrabajoDocument14 pagesEl Tratamiento de Los Medios Probatorios en La Nueva Ley Procesal Del TrabajoJorge Huamani Salinas100% (1)
- Emocionesjesus PDFDocument31 pagesEmocionesjesus PDFVictor JimenezNo ratings yet
- Teoría de Galtung ResumenDocument30 pagesTeoría de Galtung ResumenJuan LozanoNo ratings yet
- 333 Formato Guia Admitir DemandaDocument4 pages333 Formato Guia Admitir DemandaJosé Ricardo Hernández GómezNo ratings yet
- Guia #2 Sociales 11ºDocument6 pagesGuia #2 Sociales 11ºleonardo bermudezNo ratings yet
- Los Haluros de AlquiloDocument28 pagesLos Haluros de AlquiloMariana Luna HdezNo ratings yet
- Ficha de Mitosis PDFDocument2 pagesFicha de Mitosis PDFPaloma BPNo ratings yet
- Guabloche ColungeDocument1 pageGuabloche ColungeJamil Vacas BlasNo ratings yet
- Romero, José Luis - La Edad Media. Guía DidácticaDocument5 pagesRomero, José Luis - La Edad Media. Guía DidácticaVerónica Díaz PereyroNo ratings yet
- Cultura de La Paz EquipoDocument7 pagesCultura de La Paz EquipoAna BalderasNo ratings yet
- Lamentaciones de Un Prepucio Shalom Auslander PDFDocument522 pagesLamentaciones de Un Prepucio Shalom Auslander PDFMarco Aurelio Castaño AriasNo ratings yet
- Transcriptomica - Fitopatologia Molecular 2016Document78 pagesTranscriptomica - Fitopatologia Molecular 2016CAMILA PATRICIA ROJAS PIZARRONo ratings yet
- La Literatura de FronteraDocument21 pagesLa Literatura de FronterasyjacobirNo ratings yet
- Sesión de Tutoria-03-04Document7 pagesSesión de Tutoria-03-04Srocio VicencioNo ratings yet
- Teleología Contable - Aplicación de Las Ocho Ramas de La Filosofía A La ContabilidadDocument21 pagesTeleología Contable - Aplicación de Las Ocho Ramas de La Filosofía A La ContabilidadJhon MendozaNo ratings yet
- Diapositivas SemiologiaDocument17 pagesDiapositivas SemiologiaJhessica CalvoNo ratings yet
- Importar Los Archivos XLS A AUDIMSS PDFDocument84 pagesImportar Los Archivos XLS A AUDIMSS PDFEmmanuel VelaNo ratings yet
- PE 2015 Reunion TecnicaDocument40 pagesPE 2015 Reunion Tecnicanancy pichihua alataNo ratings yet
- MIELOMENINGOCELEDocument6 pagesMIELOMENINGOCELEesteba gomezNo ratings yet
- Guía para La Formulación de Un Plan de Contingencias InformáticasDocument7 pagesGuía para La Formulación de Un Plan de Contingencias InformáticasRODJAVNo ratings yet
- Caso Cerro PanteónDocument9 pagesCaso Cerro PanteónYasna SandovalNo ratings yet
- Actividad Ficha 2Document2 pagesActividad Ficha 2Sandra Navarro AguadoNo ratings yet
- 01 Cambio Linguistico - ZannierDocument34 pages01 Cambio Linguistico - Zannierstephanie_canetNo ratings yet
- Cuestionario CompiladoresDocument8 pagesCuestionario CompiladoresCarlos Meneses HernandezNo ratings yet
- Metodología de La InvestigaciónDocument6 pagesMetodología de La InvestigaciónDarwin GutierrezNo ratings yet
- Nuevos Activos AntioxidantesDocument24 pagesNuevos Activos AntioxidantesCRAULNo ratings yet
- Programacion Anual Inglés 5° Grado 2023Document14 pagesProgramacion Anual Inglés 5° Grado 2023Julio MontoyaNo ratings yet
- Biografía de León XIIIDocument4 pagesBiografía de León XIIIjarumiNo ratings yet