You might also like
- 3rd Lecture Morphological Changes During Maturation o Te GameteaDocument25 pages3rd Lecture Morphological Changes During Maturation o Te GameteaHussein Al Saedi100% (1)
- PBL SS 2 - Fetal DevelopmentDocument3 pagesPBL SS 2 - Fetal DevelopmentJellie MendozaNo ratings yet
- Textbook of Cardiovascular Medicine in Dogs and Cats (VetBooks - Ir)Document1,112 pagesTextbook of Cardiovascular Medicine in Dogs and Cats (VetBooks - Ir)Jonathan Mann100% (1)
- Neurulation Refers To The Folding Process in Vertebrate Embryos, Which Includes TheDocument6 pagesNeurulation Refers To The Folding Process in Vertebrate Embryos, Which Includes Theaaliya saaheenNo ratings yet
- GastrulationDocument1 pageGastrulationClaire BolalinNo ratings yet
- Cleavage: The Main Process of Animal DevelopmentDocument123 pagesCleavage: The Main Process of Animal DevelopmentNUR ASMA RIZKYNo ratings yet
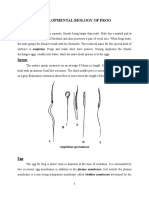
- Development of Frog Umanga Chapagain Read OnlyDocument9 pagesDevelopment of Frog Umanga Chapagain Read OnlySubarna PudasainiNo ratings yet
- Anatomy and Physiology Chapter 3Document55 pagesAnatomy and Physiology Chapter 3Yasin Putra EsbeyeNo ratings yet
- FAQ in MicrobiologyDocument117 pagesFAQ in MicrobiologyVenkatesh Arumugam100% (1)
- Developmental Biology of Frog: SpermDocument21 pagesDevelopmental Biology of Frog: SpermRavindra MadurNo ratings yet
- Fertilization &implantationDocument30 pagesFertilization &implantationAzza100% (1)
- Early DevelopmentDocument5 pagesEarly DevelopmentMarina of The SeaNo ratings yet
- Gastrulation in AmphibiansDocument20 pagesGastrulation in AmphibiansAnonymous mpD5IsJvaNo ratings yet
- 01 - Embryology (Theory)Document54 pages01 - Embryology (Theory)Simran Kashyap100% (1)
- Embryonic Period: Dr. Khin Ma MADocument34 pagesEmbryonic Period: Dr. Khin Ma MAIbrahim Foondun100% (1)
- Teratology (Birth Defect) (Birth Defect) : Lecture NoteDocument36 pagesTeratology (Birth Defect) (Birth Defect) : Lecture NoteTofik MohammedNo ratings yet
- Gametogenesis: Spermatogenesis & Oogenesis: Dr. Naveen Kumar MMMC ManipalDocument16 pagesGametogenesis: Spermatogenesis & Oogenesis: Dr. Naveen Kumar MMMC ManipalLim Yi QiNo ratings yet
- 8.introduction To EmbryologyDocument63 pages8.introduction To EmbryologyAhmed OrabyNo ratings yet
- Embryology NotesDocument23 pagesEmbryology NotesMichelle RequejoNo ratings yet
- IntroductionDocument24 pagesIntroductioninoka911No ratings yet
- Human Reroduction by Digambar Mali.Document22 pagesHuman Reroduction by Digambar Mali.Surge E-Solution Pvt.Ltd.100% (1)
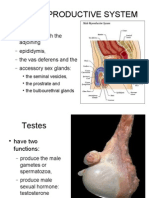
- Reproductive SystemDocument42 pagesReproductive SystemAileen Grace Delima100% (1)
- Bacteriology PDFDocument19 pagesBacteriology PDFDarshana Juvekar0% (1)
- Lecture 3 GametogenesisDocument28 pagesLecture 3 GametogenesisAparna PSNo ratings yet
- SerialDilutions PDFDocument5 pagesSerialDilutions PDFAmelia_KharismayantiNo ratings yet
- Embryology - Science That Studies The Early Development and Formation of The OrganismDocument27 pagesEmbryology - Science That Studies The Early Development and Formation of The OrganismMeah PachecoNo ratings yet
- Practical Laboratory Diagnosis of Parasitic DiseasesDocument48 pagesPractical Laboratory Diagnosis of Parasitic DiseasesliliposhianNo ratings yet
- Fetal DevelopmentDocument15 pagesFetal DevelopmentOmar HammadNo ratings yet
- Lymphoid SystemDocument11 pagesLymphoid SystemSheena PasionNo ratings yet
- Class CestodaDocument114 pagesClass CestodaICAY100% (1)
- Respiratorysystem 120105050542 Phpapp01Document43 pagesRespiratorysystem 120105050542 Phpapp01Buuvee BeNo ratings yet
- General Microbiology - Introduction - 1Document3 pagesGeneral Microbiology - Introduction - 1Harsha N KumarNo ratings yet
- Intermediate FilamentsDocument4 pagesIntermediate FilamentsSai SridharNo ratings yet
- Rubella - Pediatrics - MSD Manual Professional EditionDocument4 pagesRubella - Pediatrics - MSD Manual Professional EditionSadia Shafique100% (1)
- 07 Gastrulation BiologyDocument22 pages07 Gastrulation BiologyKashif JamalNo ratings yet
- Amniotic Fluid: Group 5Document29 pagesAmniotic Fluid: Group 5Francis ValdezNo ratings yet
- Chick NeurulationDocument14 pagesChick NeurulationSophia RubiaNo ratings yet
- 4+5 Tissue Types and Epithelial TissueDocument44 pages4+5 Tissue Types and Epithelial Tissueyasser100% (2)
- History and Branches of MicrobiologyDocument77 pagesHistory and Branches of Microbiologyrafaqat ahmedNo ratings yet
- Unit 1 MicrobiologyDocument7 pagesUnit 1 MicrobiologySunil SharmaNo ratings yet
- Early Development in BirdsDocument53 pagesEarly Development in BirdsMuqaddas NadeemNo ratings yet
- Female Reproductive HistologyDocument59 pagesFemale Reproductive HistologyIta Indriani100% (2)
- Early Embryonic DevelopmentDocument19 pagesEarly Embryonic DevelopmentderhangkerNo ratings yet
- Cell and TissuesDocument18 pagesCell and Tissuesalvin jr ayugatNo ratings yet
- SpermatogenesisDocument24 pagesSpermatogenesisOmm KoliNo ratings yet
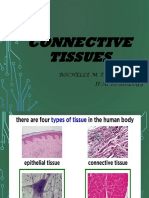
- Connective Tissues: Rochelle M. Delos Santos Ii-A1 Bs-BiologyDocument54 pagesConnective Tissues: Rochelle M. Delos Santos Ii-A1 Bs-BiologyJordan Ramos100% (1)
- 4th To 8th WeekDocument48 pages4th To 8th WeekjefferyNo ratings yet
- Developmental Biology, 12th Edition (Michael J.F. Barresi, Scott F. Gilbert)Document48 pagesDevelopmental Biology, 12th Edition (Michael J.F. Barresi, Scott F. Gilbert)Nita DelinaNo ratings yet
- Human Embryonic DevelopmentDocument8 pagesHuman Embryonic DevelopmentGargi BhattacharyaNo ratings yet
- Appropriate Clinical Specimens Collection and Transport For Diagnostic VirologyDocument48 pagesAppropriate Clinical Specimens Collection and Transport For Diagnostic Virologylong thomNo ratings yet
- Kaufman’s Atlas of Mouse Development Supplement: With Coronal SectionsFrom EverandKaufman’s Atlas of Mouse Development Supplement: With Coronal SectionsRichard BaldockNo ratings yet
- Phinma - University of PangasinanDocument2 pagesPhinma - University of PangasinanRonnie De Vera IINo ratings yet
- Female Reproductive System: UterusDocument44 pagesFemale Reproductive System: UterusBeni KurniawanNo ratings yet
- Micrology and Parasitology ReviewerDocument44 pagesMicrology and Parasitology Reviewermarjourie cadampog100% (1)
- Overview of The Immune SystemDocument9 pagesOverview of The Immune SystemAyadi P. AkinwandeNo ratings yet
- MCN LectureDocument8 pagesMCN LectureEmily BernatNo ratings yet
- Reproductive PhysiologyDocument40 pagesReproductive PhysiologyBaiq Trisna Satriana100% (1)
- 01 OogenesisDocument21 pages01 OogenesisMu LokNo ratings yet
- Histology of BreastDocument2 pagesHistology of BreastIbrahim SugiyonoNo ratings yet
- Man Meets Microbes: An Introduction to Medical MicrobiologyFrom EverandMan Meets Microbes: An Introduction to Medical MicrobiologyNo ratings yet
- Antithyroid DrugsDocument52 pagesAntithyroid DrugsYeyen MusainiNo ratings yet
- Basics of Cytogenetics: Mitosis and Meiosis TutorialDocument17 pagesBasics of Cytogenetics: Mitosis and Meiosis TutorialYeyen MusainiNo ratings yet
- Principles of Dna Isolation & PurificationDocument47 pagesPrinciples of Dna Isolation & PurificationYeyen Musaini100% (1)
- Outcome - A Prospective Study in A Third-World CountryDocument2 pagesOutcome - A Prospective Study in A Third-World CountryYeyen MusainiNo ratings yet
- Development of FrogDocument44 pagesDevelopment of Frogsuyog raj gautamNo ratings yet
- A Laboratory Manual of Vertebrate Embryology A Laboratory Manual of Vertebrate EmbryologyDocument96 pagesA Laboratory Manual of Vertebrate Embryology A Laboratory Manual of Vertebrate EmbryologyANNE MARGUERITE BARETENo ratings yet
- Worksheet For Morgan - Carter Laboratory #25 "Animal Development"Document9 pagesWorksheet For Morgan - Carter Laboratory #25 "Animal Development"HyenaNo ratings yet
- Practical MCQ Sheet - QuestionsDocument6 pagesPractical MCQ Sheet - QuestionsIoana SavaNo ratings yet
- Fate Mapping - Gastrulation - Lec Notes - KMZRDocument27 pagesFate Mapping - Gastrulation - Lec Notes - KMZRSaad SakalNo ratings yet
- 05 GastrulationDocument6 pages05 GastrulationcarlgangcaNo ratings yet
- Embryologydevelopmental AnatomyDocument62 pagesEmbryologydevelopmental AnatomygirishNo ratings yet
- Biology 453 - Comparative Vert. Anatomy WEEK 1, LAB 2: Embryology of Frog & ChickDocument9 pagesBiology 453 - Comparative Vert. Anatomy WEEK 1, LAB 2: Embryology of Frog & ChickPunk Midget-FairyNo ratings yet
- Developmental Biology QuestionsDocument10 pagesDevelopmental Biology QuestionsSurajit Bhattacharjee100% (1)
- Early Development in BirdsDocument53 pagesEarly Development in BirdsMuqaddas NadeemNo ratings yet
- Practical - DEVELOPMENTAL STAGES OF FROGDocument6 pagesPractical - DEVELOPMENTAL STAGES OF FROGtooba tahirNo ratings yet
- Developmental Biology of Frog: SpermDocument21 pagesDevelopmental Biology of Frog: SpermRavindra MadurNo ratings yet
- Principles of Development: Developmental BiologyDocument86 pagesPrinciples of Development: Developmental BiologyPaolo OcampoNo ratings yet
- General Stem Cell Biology BrochureDocument32 pagesGeneral Stem Cell Biology BrochureSigma-AldrichNo ratings yet
- Development of AmphioxusDocument59 pagesDevelopment of AmphioxusDyah Ayu PuspitasariNo ratings yet
- Obero, Jecamiah E. - LAB # 6Document5 pagesObero, Jecamiah E. - LAB # 6JECAMIAH OBERONo ratings yet
- EmbryologyDocument68 pagesEmbryologyBj Larracas100% (1)
- Developmental Biology Exam 1 Quiz 1 2 CH 13 Online Book Drosophila Online BookDocument10 pagesDevelopmental Biology Exam 1 Quiz 1 2 CH 13 Online Book Drosophila Online BookPearl PascuaNo ratings yet
- Zoology 2Document16 pagesZoology 2Amod KumarNo ratings yet
- Book - The Frog Its Reproduction and Development 17 - EmbryologyDocument12 pagesBook - The Frog Its Reproduction and Development 17 - EmbryologyJudy Anne Vitales GalizaNo ratings yet
- Principles of Embryonic DevelopmentDocument29 pagesPrinciples of Embryonic DevelopmentMihailo Majk ŽikićNo ratings yet
- LSM3233 Lecture 2 Notes 2013Document44 pagesLSM3233 Lecture 2 Notes 2013Prab NathanNo ratings yet
- Active Gel Physics: J. Prost, F. Jülicher and J-F. JoannyDocument7 pagesActive Gel Physics: J. Prost, F. Jülicher and J-F. JoannyElviaNo ratings yet
- 24 Hour Chick Embryo - Embryology LabDocument3 pages24 Hour Chick Embryo - Embryology LabIvy Cruz50% (2)
- Karla Loureiro Almeida, José Abreu, C. Y. Irene Yan (Auth.), Henning Ulrich (Eds.)-Perspectives of Stem Cells_ From Tools for Studying Mechanisms of Neuronal Differentiation Towards Therapy-Springer NDocument300 pagesKarla Loureiro Almeida, José Abreu, C. Y. Irene Yan (Auth.), Henning Ulrich (Eds.)-Perspectives of Stem Cells_ From Tools for Studying Mechanisms of Neuronal Differentiation Towards Therapy-Springer NPhilippeMeirelles100% (1)
- Biology 453 - Comparative Vert. Anatomy WEEK 1, LAB 2: Embryology of Frog & ChickDocument6 pagesBiology 453 - Comparative Vert. Anatomy WEEK 1, LAB 2: Embryology of Frog & ChickMuhammad WildanulhaqNo ratings yet
- 3-Kingdom AnimaliaDocument16 pages3-Kingdom AnimaliaEhmz XavièrNo ratings yet
- Embryology ObgDocument36 pagesEmbryology ObgsamrusangaliNo ratings yet
- Gosner 1960 - A Simplified Table For Staging Anuran Embryos and Larvae With Notes On IdentificationDocument9 pagesGosner 1960 - A Simplified Table For Staging Anuran Embryos and Larvae With Notes On IdentificationRuivo LucasNo ratings yet