You might also like
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (121)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (400)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- Workbook3 ExcretionDocument12 pagesWorkbook3 ExcretionDarambazar GantulgaNo ratings yet
- Head and Neck Cheat Sheets PDFDocument8 pagesHead and Neck Cheat Sheets PDFJM LiensNo ratings yet
- The Sea Urchin Fertilization LabDocument5 pagesThe Sea Urchin Fertilization LabAmy HollingsworthNo ratings yet
- 5 - Cluster 1 - Maintaining A Healthy Body - Manitoba - Grade 5 Science UnitDocument75 pages5 - Cluster 1 - Maintaining A Healthy Body - Manitoba - Grade 5 Science UnitZabrinaNo ratings yet
- Noninfectious Diseases and Pathology of Reptiles, Color Atlas and Text, 2nd Edition (VetBooks - Ir)Document536 pagesNoninfectious Diseases and Pathology of Reptiles, Color Atlas and Text, 2nd Edition (VetBooks - Ir)Letícia de Oliveira SousaNo ratings yet
- Pulmonary EmbolismDocument23 pagesPulmonary EmbolismBianca Dizon0% (1)
- Phaeophyta (Brown Algae)Document22 pagesPhaeophyta (Brown Algae)Sona PectusNo ratings yet
- Practicum 1Document4 pagesPracticum 1armin509No ratings yet
- Electrochemistry: NotesDocument14 pagesElectrochemistry: Notesarmin509No ratings yet
- Carbohydrate Metabolism: NotesDocument15 pagesCarbohydrate Metabolism: Notesarmin509No ratings yet
- Lesson 19Document6 pagesLesson 19armin509No ratings yet
- Assignment QualityDocument1 pageAssignment Qualityarmin509No ratings yet
- Lesson 10Document13 pagesLesson 10armin509No ratings yet
- Basic Pathology Repair. Cell Growth and Regeneration. Wound HealingDocument4 pagesBasic Pathology Repair. Cell Growth and Regeneration. Wound Healingarmin509No ratings yet
- Basic Pathology Chapter 11: Artherosclerosis: A. How It StartsDocument2 pagesBasic Pathology Chapter 11: Artherosclerosis: A. How It Startsarmin509No ratings yet
- Unit 3, Microscope PresentationDocument33 pagesUnit 3, Microscope Presentationarmin509No ratings yet
- ABS Cell PathologyDocument26 pagesABS Cell Pathologyarmin509No ratings yet
- Microscope PresentationDocument16 pagesMicroscope Presentationarmin509No ratings yet
- Communication Process SalesmanshipDocument18 pagesCommunication Process SalesmanshipCrissa Marcelo MarianoNo ratings yet
- External Parts of The Humans BodyDocument8 pagesExternal Parts of The Humans BodyDian kusriniNo ratings yet
- Week 3 - Muscles LabDocument19 pagesWeek 3 - Muscles LabMarx AsuncionNo ratings yet
- REPRODUCTION DEVELOPMENT - PRACTICE QUESTIONS-Student Copy3Document16 pagesREPRODUCTION DEVELOPMENT - PRACTICE QUESTIONS-Student Copy3salinaNo ratings yet
- Physiology One LinersDocument10 pagesPhysiology One LinersadiNo ratings yet
- Anatomy and Physiology of The Respiratory SystemDocument9 pagesAnatomy and Physiology of The Respiratory SystemMitchy SunNo ratings yet
- The Internal and External Anatomy of FishDocument5 pagesThe Internal and External Anatomy of FishGienelle BermidoNo ratings yet
- PyelonephritisDocument59 pagesPyelonephritisGheyl GabrielNo ratings yet
- Orca Share Media1673167717323 7017774049441689006Document53 pagesOrca Share Media1673167717323 7017774049441689006Rosenifah CairodingNo ratings yet
- Non Hodgkin Lymphoma PresentationDocument17 pagesNon Hodgkin Lymphoma PresentationClayton Green100% (1)
- Pulmonary Embolism Is A Common and Potentially Lethal ConditionDocument13 pagesPulmonary Embolism Is A Common and Potentially Lethal ConditionJasleen KaurNo ratings yet
- Iicp 1Document5 pagesIicp 1Melodia Turqueza GandezaNo ratings yet
- LEOPOLD MANEUVER RetdemDocument3 pagesLEOPOLD MANEUVER Retdemjohncarlo ramosNo ratings yet
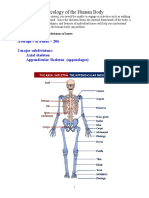
- Osteology of The Human BodyDocument17 pagesOsteology of The Human BodyAyuba EmmanuelNo ratings yet
- MATERNAL SERUM ALFA FETO PROTEIN (Jyoti Singh)Document11 pagesMATERNAL SERUM ALFA FETO PROTEIN (Jyoti Singh)Jyothi Singh SuryavanshiNo ratings yet
- 2ND Periodical Test - Science 5Document6 pages2ND Periodical Test - Science 5Eugel Garedo50% (2)
- Biology ProjectDocument21 pagesBiology ProjectBad Time?85% (13)
- HeartDocument12 pagesHeartRebar photographyNo ratings yet
- 1 Gcse Aqa BiologyDocument43 pages1 Gcse Aqa Biologycharlotteamy10No ratings yet
- Prenatal Development Part 1 by Lea CortezDocument26 pagesPrenatal Development Part 1 by Lea CortezLea Cabillo CortezNo ratings yet
- Nutrients and Waste Transport SystemDocument22 pagesNutrients and Waste Transport SystemItsPotatoQueen MaeNo ratings yet
- Romanowsky Stain - WikipediaDocument1 pageRomanowsky Stain - WikipediaANo ratings yet
- 1-Human Physiology Digestion Absorption PDFDocument29 pages1-Human Physiology Digestion Absorption PDFPintu KarmakarNo ratings yet
- PPT1.2 Cell Doctrine PDFDocument44 pagesPPT1.2 Cell Doctrine PDFBharath KumarNo ratings yet