You might also like
- Current Concepts of ImmunoprophylaxisDocument31 pagesCurrent Concepts of ImmunoprophylaxisTina Ong SinagaNo ratings yet
- Pet-to-Man Travelling Staphylococci: A World in ProgressFrom EverandPet-to-Man Travelling Staphylococci: A World in ProgressVincenzo SaviniNo ratings yet
- Cultivation of VirusesDocument34 pagesCultivation of VirusesYogesh BanginwarNo ratings yet
- SARS-CoV-2 Viral Outbreak Investigation: Laboratory Perspective: Clinical Updates in COVID-19From EverandSARS-CoV-2 Viral Outbreak Investigation: Laboratory Perspective: Clinical Updates in COVID-19Rating: 3 out of 5 stars3/5 (1)
- Vaccine: - Chhabi Acharya Binod Rasaili Rupesh K.C Hope Int'l CollegeDocument21 pagesVaccine: - Chhabi Acharya Binod Rasaili Rupesh K.C Hope Int'l CollegeScott Domes100% (1)
- Bacterial VaccineDocument44 pagesBacterial VaccineShrenik LuniyaNo ratings yet
- Scope of MicroDocument32 pagesScope of MicroShehram QazafiNo ratings yet
- Cultivation of Viruses: Embryonated EggDocument26 pagesCultivation of Viruses: Embryonated EggMubashir AzizNo ratings yet
- ZIKA VIRUS INFECTION Emerging Public Health ConcernDocument36 pagesZIKA VIRUS INFECTION Emerging Public Health Concerntummalapalli venkateswara rao100% (1)
- DR Channa Senanayake Dept of MicrobiologyDocument19 pagesDR Channa Senanayake Dept of MicrobiologyTharinduNo ratings yet
- Immunology - Chapter 19 VaccinesDocument21 pagesImmunology - Chapter 19 VaccinesJJHHJHi100% (1)
- COVID Vaccine Systematic Review Dong Dai WeiDocument14 pagesCOVID Vaccine Systematic Review Dong Dai WeiJason SnapeNo ratings yet
- 01 - General Feature of VirusDocument2 pages01 - General Feature of VirusSajid Ali100% (1)
- By Meenakshi Bohra M.SC Biotech (Prev.)Document24 pagesBy Meenakshi Bohra M.SC Biotech (Prev.)motional100% (1)
- Vaccine Production (L1F17PHMD0174)Document22 pagesVaccine Production (L1F17PHMD0174)Faizah Khalid100% (1)
- Chapter 10 Environmental and Applied MicrobiologyDocument44 pagesChapter 10 Environmental and Applied MicrobiologySherinne Jane CariazoNo ratings yet
- Research-Diagnosis Knowledge UnderstandingDocument29 pagesResearch-Diagnosis Knowledge Understandingnvd280189No ratings yet
- Vaccine & Principle of ImmunizationDocument23 pagesVaccine & Principle of ImmunizationSindhu Babu100% (1)
- Bacterial Genetics & ReproductionDocument18 pagesBacterial Genetics & ReproductionAsimNo ratings yet
- Viral ReplicationDocument1 pageViral ReplicationMoh'd Ghanayem100% (1)
- Microbial Genetics: Julia Hartati, DRDocument21 pagesMicrobial Genetics: Julia Hartati, DRerwan_mivecNo ratings yet
- Cultivation of VirusDocument45 pagesCultivation of VirusPRANAB DEYNo ratings yet
- Viral ReplicationDocument26 pagesViral ReplicationJhann100% (1)
- Vaccine Development, Testing, and RegulationDocument12 pagesVaccine Development, Testing, and RegulationP Bijoya SinghaNo ratings yet
- General MycologyDocument28 pagesGeneral MycologyShimelis Teshome AyalnehNo ratings yet
- Appropriate Clinical Specimens Collection and Transport For Diagnostic VirologyDocument48 pagesAppropriate Clinical Specimens Collection and Transport For Diagnostic Virologylong thomNo ratings yet
- Phylogenetic TreesDocument20 pagesPhylogenetic TreesRamesh Kandagatla100% (1)
- Edible VaccinesDocument21 pagesEdible VaccinesPritish SareenNo ratings yet
- New Developments in Chromogenic and Fluorogenic Culture MediaDocument14 pagesNew Developments in Chromogenic and Fluorogenic Culture MediaPaulina LizzethNo ratings yet
- Alternative Culture Media For Bacterial Growth Using Different Formulation of Protein SourcesDocument4 pagesAlternative Culture Media For Bacterial Growth Using Different Formulation of Protein SourcesMasitha Dwi AmiraNo ratings yet
- Classification of VirusesDocument2 pagesClassification of Viruseshely shahNo ratings yet
- Viral DiagnosisDocument37 pagesViral Diagnosissalamshakir56No ratings yet
- Bot 101 NotesDocument2 pagesBot 101 NotesAbid Hussain100% (1)
- Virus CultivationDocument33 pagesVirus Cultivationved.g007No ratings yet
- Viruses: RNA Virus Reverse Genetics and Vaccine DesignDocument20 pagesViruses: RNA Virus Reverse Genetics and Vaccine Designfragariavesca100% (1)
- Module 4 Annex Part 1 MKDocument95 pagesModule 4 Annex Part 1 MKtata_77No ratings yet
- Microbial Diseases of The Skin and EyesDocument29 pagesMicrobial Diseases of The Skin and EyesqueennieNo ratings yet
- Norovirus ESSAYDocument4 pagesNorovirus ESSAYrasharagheiiNo ratings yet
- Vaccines PART 1Document57 pagesVaccines PART 1shahbaz100% (1)
- Recent Trends in Viral VaccinesDocument64 pagesRecent Trends in Viral VaccinesAkula Sai PrasannaNo ratings yet
- Pathogenic SpirochetesDocument45 pagesPathogenic SpirochetesManisanthosh Kumar100% (1)
- General Characteristics of Viruses:: VirologyDocument12 pagesGeneral Characteristics of Viruses:: VirologyJamine Joyce Ortega-AlvarezNo ratings yet
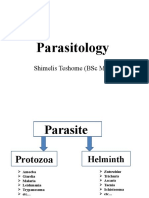
- ParasitologyDocument107 pagesParasitologyShimelis Teshome AyalnehNo ratings yet
- Diagnosis of VirusesDocument21 pagesDiagnosis of VirusesOlivia Wesula Lwande DanielleNo ratings yet
- Validation of Viral Clearance: S K Patel College of Pharmaceutical Education & Research Ganpat UniversityDocument24 pagesValidation of Viral Clearance: S K Patel College of Pharmaceutical Education & Research Ganpat UniversitynirmalpharmaNo ratings yet
- Cultivation of VirusDocument26 pagesCultivation of VirusMariam0% (1)
- Specific Host Defenses: The Immune ResponseDocument54 pagesSpecific Host Defenses: The Immune Responseadyaly44No ratings yet
- Branches of MicrobiologyDocument2 pagesBranches of Microbiologyhaseeb_khan321294067% (3)
- Smallpox MonkeypoxDocument63 pagesSmallpox MonkeypoxjufhiurhNo ratings yet
- Introduction To: ParasitesDocument32 pagesIntroduction To: ParasitesJaznMonNo ratings yet
- Mark Old Corne Microbiological ValidationDocument28 pagesMark Old Corne Microbiological ValidationhumusdelombrizNo ratings yet
- Laboratory Diagnosis of Viral InfectionDocument37 pagesLaboratory Diagnosis of Viral InfectionstudymedicNo ratings yet
- Core Topic 4 - The Different Types of Vaccines Used and Their Composition - March 2009Document27 pagesCore Topic 4 - The Different Types of Vaccines Used and Their Composition - March 2009Pranav NakhateNo ratings yet
- PH Antimicrobial: Ms. Sunisa Thongdee 4518996 Ms - Janjira Sillapee 4518999Document19 pagesPH Antimicrobial: Ms. Sunisa Thongdee 4518996 Ms - Janjira Sillapee 4518999biochemi100% (2)
- Human PapillomavirusDocument5 pagesHuman PapillomavirusSofiaLopezNo ratings yet
- Animal Cell CultureDocument34 pagesAnimal Cell CultureCata NaviaNo ratings yet
- A Review of Foot-And-Mouth Disease With Special Consideration For The Clinical and Epidemiological FactorsDocument13 pagesA Review of Foot-And-Mouth Disease With Special Consideration For The Clinical and Epidemiological FactorsjaherrerarNo ratings yet
- Human Immunodeficiency Virus InfectionDocument20 pagesHuman Immunodeficiency Virus InfectionFatos ShuliNo ratings yet
- 6476 Formulation of Alternative Culture Media For Bacterial and Fungal GrowthDocument6 pages6476 Formulation of Alternative Culture Media For Bacterial and Fungal GrowthHanief MitsukhaNo ratings yet
- Can Marijuana Market5Document21 pagesCan Marijuana Market5SyedNo ratings yet
- PB India Snapshot March2018Document4 pagesPB India Snapshot March2018SyedNo ratings yet
- 2015 Pharma and Bioteh PreviewDocument12 pages2015 Pharma and Bioteh PreviewSyedNo ratings yet
- HRS Funding Opportunities BulletinSeptember222017Document24 pagesHRS Funding Opportunities BulletinSeptember222017SyedNo ratings yet
- PB ReviewIndiaMarch2018 PDFDocument47 pagesPB ReviewIndiaMarch2018 PDFSyedNo ratings yet
- Orphan Drug Evaluate Pharma EPOD17Document26 pagesOrphan Drug Evaluate Pharma EPOD17Uzair Ul GhaniNo ratings yet
- 2015 Pharma and Bioteh Preview PDFDocument12 pages2015 Pharma and Bioteh Preview PDFSyedNo ratings yet
- 1 Presence of Indian Pharma in USMkt 29 Jun 2009Document12 pages1 Presence of Indian Pharma in USMkt 29 Jun 2009SyedNo ratings yet
- 2012 Indian-Pharma-Sector PDFDocument50 pages2012 Indian-Pharma-Sector PDFSyedNo ratings yet
- Case Study DR Reddy-Industry PDFDocument61 pagesCase Study DR Reddy-Industry PDFSyed0% (2)
- WHO Units in IndiaDocument45 pagesWHO Units in IndiaSyed100% (1)
- India's Hospital EcosystemDocument7 pagesIndia's Hospital EcosystemSyedNo ratings yet
- Nutraceutical Products in IndiaDocument7 pagesNutraceutical Products in IndiaSyedNo ratings yet
- 30 Daysto Where You Wantto BeDocument44 pages30 Daysto Where You Wantto BeAlexandra Ene100% (2)
- 5 Biotechs To Watch TaiwanDocument18 pages5 Biotechs To Watch TaiwanSyedNo ratings yet
- BSum Behind The Cloud PDFDocument9 pagesBSum Behind The Cloud PDFSyedNo ratings yet
- 50 Effective Email PDFDocument123 pages50 Effective Email PDFSyedNo ratings yet
- WHO Phleb - Manual - Ready PDFDocument127 pagesWHO Phleb - Manual - Ready PDFSyedNo ratings yet
- Characteristics of Life UpdatedDocument12 pagesCharacteristics of Life UpdatedYla Mae Buo AcejoNo ratings yet
- 10 - Enzymes - PPT - AutoRecoveredDocument65 pages10 - Enzymes - PPT - AutoRecoveredFaith WilliamsNo ratings yet
- Ijms 23 03754Document14 pagesIjms 23 03754Benyam ZenebeNo ratings yet
- Boon or BaneDocument5 pagesBoon or BanesujeeNo ratings yet
- Growth and Development PDFDocument12 pagesGrowth and Development PDFAkshayNo ratings yet
- Aakash Botany Study Package 4 SolutionssssDocument120 pagesAakash Botany Study Package 4 SolutionssssRishika PaulNo ratings yet
- BT503 - Past Papers PDFDocument83 pagesBT503 - Past Papers PDFJawad100% (1)
- SDS PageDocument12 pagesSDS PageShadia HeyariNo ratings yet
- Hassan Et Al., 2024Document15 pagesHassan Et Al., 2024Herda CahyaningrumNo ratings yet
- Importance of Weak ABO SubgroupsDocument3 pagesImportance of Weak ABO SubgroupsCharmaine Corpuz GranilNo ratings yet
- CHAPTER 16 MicroparaDocument46 pagesCHAPTER 16 MicroparaKathlyn LopeñaNo ratings yet
- BIOLOGY Form 5 Chapter 1 TransportDocument42 pagesBIOLOGY Form 5 Chapter 1 TransportShephard Png91% (95)
- Mutagenesis Application GuideDocument57 pagesMutagenesis Application GuideCesa Mulyo100% (1)
- Retinoid Receptors and Therapeutic Applications of RAR/RXR ModulatorsDocument23 pagesRetinoid Receptors and Therapeutic Applications of RAR/RXR ModulatorsAKellyNMNo ratings yet
- Evolution-History and EvidenceDocument19 pagesEvolution-History and Evidencekatexxx100% (1)
- The Digestive System MODULEDocument3 pagesThe Digestive System MODULEBen Moldenhaur100% (2)
- A Method For Turbidimetric Measurement of BacterialDocument4 pagesA Method For Turbidimetric Measurement of BacterialCrisMayaNo ratings yet
- Olive Leaf Extract (HIV Fighter, Yeast Buster, Viral Killer)Document16 pagesOlive Leaf Extract (HIV Fighter, Yeast Buster, Viral Killer)XR500Final100% (8)
- Buenasher Learning Academy Inc.: Heredity: Inheritance and VariationDocument4 pagesBuenasher Learning Academy Inc.: Heredity: Inheritance and VariationMaam Elle CruzNo ratings yet
- 9700 Y16 Sy 1Document81 pages9700 Y16 Sy 1api-255467062No ratings yet
- Statement of PurposeDocument2 pagesStatement of PurposeRobert Daniel95% (21)
- CAMC 102 - General Physiology and ToxicologyDocument6 pagesCAMC 102 - General Physiology and ToxicologyPrincess Mae AspuriaNo ratings yet
- Anatomi TumbuhanDocument30 pagesAnatomi TumbuhanMEISYI DSNo ratings yet
- Definisi Stress 1Document30 pagesDefinisi Stress 1chatarinaNo ratings yet
- Brs Embryology Q&ADocument41 pagesBrs Embryology Q&ASURG BLOCK100% (1)
- Virus Introduction of Virus: Martinus Beijerinck Discovered The Tobacco Mosaic Virus (TMV) From TheDocument5 pagesVirus Introduction of Virus: Martinus Beijerinck Discovered The Tobacco Mosaic Virus (TMV) From Therafiul muttakimNo ratings yet
- A Novel in Vitro Hydroponic Culture System For PotatoDocument5 pagesA Novel in Vitro Hydroponic Culture System For PotatoPilar7100No ratings yet
- Worksheet 9.1: Chapter 9: Plant Science - Fifteen Summary FactsDocument2 pagesWorksheet 9.1: Chapter 9: Plant Science - Fifteen Summary FactsRabia RafiqueNo ratings yet
- Alzheimers-Disease-Genetics-Fact-Sheet 0Document8 pagesAlzheimers-Disease-Genetics-Fact-Sheet 0api-285676076No ratings yet
- Biochem Case Problems 02Document4 pagesBiochem Case Problems 02karthikskamathNo ratings yet
- Uncontrolled Spread: Why COVID-19 Crushed Us and How We Can Defeat the Next PandemicFrom EverandUncontrolled Spread: Why COVID-19 Crushed Us and How We Can Defeat the Next PandemicNo ratings yet
- The Wisdom of Plagues: Lessons from 25 Years of Covering PandemicsFrom EverandThe Wisdom of Plagues: Lessons from 25 Years of Covering PandemicsRating: 4.5 out of 5 stars4.5/5 (6)
- Deaths of Despair and the Future of CapitalismFrom EverandDeaths of Despair and the Future of CapitalismRating: 4.5 out of 5 stars4.5/5 (30)
- Clean: Overcoming Addiction and Ending America’s Greatest TragedyFrom EverandClean: Overcoming Addiction and Ending America’s Greatest TragedyRating: 4 out of 5 stars4/5 (18)
- Summary: The Myth of Normal: Trauma, Illness, and Healing in a Toxic Culture By Gabor Maté MD & Daniel Maté: Key Takeaways, Summary & AnalysisFrom EverandSummary: The Myth of Normal: Trauma, Illness, and Healing in a Toxic Culture By Gabor Maté MD & Daniel Maté: Key Takeaways, Summary & AnalysisRating: 4 out of 5 stars4/5 (9)
- Do You Believe in Magic?: The Sense and Nonsense of Alternative MedicineFrom EverandDo You Believe in Magic?: The Sense and Nonsense of Alternative MedicineNo ratings yet
- There Are No Accidents: The Deadly Rise of Injury and Disaster—Who Profits and Who Pays the PriceFrom EverandThere Are No Accidents: The Deadly Rise of Injury and Disaster—Who Profits and Who Pays the PriceRating: 4.5 out of 5 stars4.5/5 (15)
- The Bodies of Others: The New Authoritarians, COVID-19 and The War Against the HumanFrom EverandThe Bodies of Others: The New Authoritarians, COVID-19 and The War Against the HumanRating: 4.5 out of 5 stars4.5/5 (12)
- There Are No Accidents: The Deadly Rise of Injury and Disaster—Who Profits and Who Pays the PriceFrom EverandThere Are No Accidents: The Deadly Rise of Injury and Disaster—Who Profits and Who Pays the PriceRating: 4.5 out of 5 stars4.5/5 (11)
- The Price of Health: The Modern Pharmaceutical Industry and the Betrayal of a History of CareFrom EverandThe Price of Health: The Modern Pharmaceutical Industry and the Betrayal of a History of CareRating: 4.5 out of 5 stars4.5/5 (3)
- Epic Measures: One Doctor. Seven Billion Patients.From EverandEpic Measures: One Doctor. Seven Billion Patients.Rating: 4 out of 5 stars4/5 (13)
- Mama Might Be Better Off Dead: The Failure of Health Care in Urban AmericaFrom EverandMama Might Be Better Off Dead: The Failure of Health Care in Urban AmericaNo ratings yet
- The Atlas of Disease: Mapping Deadly Epidemics and Contagion from the Plague to the CoronavirusFrom EverandThe Atlas of Disease: Mapping Deadly Epidemics and Contagion from the Plague to the CoronavirusRating: 4.5 out of 5 stars4.5/5 (10)
- Coronary: A True Story of Medicine Gone AwryFrom EverandCoronary: A True Story of Medicine Gone AwryRating: 5 out of 5 stars5/5 (4)
- Healthy Buildings: How Indoor Spaces Drive Performance and ProductivityFrom EverandHealthy Buildings: How Indoor Spaces Drive Performance and ProductivityRating: 5 out of 5 stars5/5 (2)
- Heat Wave: A Social Autopsy of Disaster in ChicagoFrom EverandHeat Wave: A Social Autopsy of Disaster in ChicagoRating: 4 out of 5 stars4/5 (40)
- Fatal Conveniences: The Toxic Products and Harmful Habits That Are Making You Sick—and the Simple Changes That Will Save Your HealthFrom EverandFatal Conveniences: The Toxic Products and Harmful Habits That Are Making You Sick—and the Simple Changes That Will Save Your HealthRating: 4 out of 5 stars4/5 (7)
- Inflamed: Deep Medicine and the Anatomy of InjusticeFrom EverandInflamed: Deep Medicine and the Anatomy of InjusticeRating: 5 out of 5 stars5/5 (12)
- The Invisible Rainbow: A History of Electricity and LifeFrom EverandThe Invisible Rainbow: A History of Electricity and LifeRating: 4.5 out of 5 stars4.5/5 (21)
- COVID-19: The Victims, The Heroes, The Comlicit, and Our New NormalFrom EverandCOVID-19: The Victims, The Heroes, The Comlicit, and Our New NormalNo ratings yet
- The Narrative Gym for Medicine: Introducing the ABT Framework for Medical Communication with Peers, Professionals, and the PublicFrom EverandThe Narrative Gym for Medicine: Introducing the ABT Framework for Medical Communication with Peers, Professionals, and the PublicNo ratings yet
- The Wuhan Cover-Up: And the Terrifying Bioweapons Arms RaceFrom EverandThe Wuhan Cover-Up: And the Terrifying Bioweapons Arms RaceNo ratings yet
- The Hair Color Mix Book: More Than 150 Recipes for Salon-Perfect Color at HomeFrom EverandThe Hair Color Mix Book: More Than 150 Recipes for Salon-Perfect Color at HomeRating: 3.5 out of 5 stars3.5/5 (7)
- Vaccine Injuries: Documented Adverse Reactions to VaccinesFrom EverandVaccine Injuries: Documented Adverse Reactions to VaccinesRating: 2.5 out of 5 stars2.5/5 (3)
- Beyond the Next Village: A Year of Magic and Medicine in NepalFrom EverandBeyond the Next Village: A Year of Magic and Medicine in NepalNo ratings yet