You might also like
- CHAPTER 22 Gluconeogenesis, Glycogen Metabolism, and The Pentose Phosphate PathwayDocument12 pagesCHAPTER 22 Gluconeogenesis, Glycogen Metabolism, and The Pentose Phosphate Pathway楊畯凱No ratings yet
- 2.2 Carbohydrate Metabolism Part 2Document13 pages2.2 Carbohydrate Metabolism Part 2John Louis PulidoNo ratings yet
- 13 - GluconeogenesisDocument23 pages13 - GluconeogenesischeckmateNo ratings yet
- Biochemistry-Exam 2 Heinz SchwarzkopfDocument162 pagesBiochemistry-Exam 2 Heinz SchwarzkopfthomasNo ratings yet
- Hexose Mono Phosphate (HMP) ShuntDocument28 pagesHexose Mono Phosphate (HMP) ShuntVishesh JainNo ratings yet
- Biochem HomeworkDocument13 pagesBiochem Homeworkfcukingfranztastik50% (2)
- Gluconeogenesis Metabolic PathwayDocument36 pagesGluconeogenesis Metabolic PathwayDayledaniel Sorveto0% (1)
- 4 Bioenergetics and Oxidative Metabolism IIDocument3 pages4 Bioenergetics and Oxidative Metabolism IILinus LiuNo ratings yet
- Chapter 20 Electron Transport and Oxidative PhosphorylationDocument14 pagesChapter 20 Electron Transport and Oxidative PhosphorylationRaabia Ansari100% (1)
- Carbohydrates SummaryDocument9 pagesCarbohydrates SummaryHarold NagunaNo ratings yet
- Biochemistry: Glycogen MetabolismDocument79 pagesBiochemistry: Glycogen Metabolismisabellacastro18No ratings yet
- Biochemistry TestDocument2 pagesBiochemistry Testbrijeshandfamily100% (1)
- Chapter 24 Practice QuestionsDocument7 pagesChapter 24 Practice QuestionsArlene F. Montalbo100% (1)
- Biochem Questions SlidesDocument46 pagesBiochem Questions SlidesTelesha Zabie50% (2)
- Biochem QbankDocument16 pagesBiochem Qbank786waqar786No ratings yet
- HW - Carbohydrate Metabolism II & Lipid MetabolismDocument2 pagesHW - Carbohydrate Metabolism II & Lipid MetabolismyanNo ratings yet
- Some Answer of Problemset - 7 - KEYDocument3 pagesSome Answer of Problemset - 7 - KEYNihir PatelNo ratings yet
- Digestion and Absorption of Dietary LipidsDocument35 pagesDigestion and Absorption of Dietary LipidsCharmaine Luc100% (2)
- Hexose Monophosphate ShuntDocument18 pagesHexose Monophosphate ShuntAbdul Jabbar Abdul JabbarNo ratings yet
- Metabolism Lippincott's Q&ADocument86 pagesMetabolism Lippincott's Q&Alina amjadNo ratings yet
- S13.CHO .MetabolismDocument18 pagesS13.CHO .MetabolismGhea Jovita SinagaNo ratings yet
- Glucose, Part1Document33 pagesGlucose, Part1SarahNo ratings yet
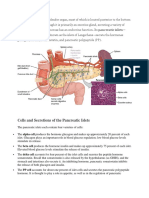
- Cells and Secretions of The Pancreatic IsletsDocument4 pagesCells and Secretions of The Pancreatic IsletsSophia OcayNo ratings yet
- PROTEIN METABOLISM Dea Farha Fira Darson FineDocument44 pagesPROTEIN METABOLISM Dea Farha Fira Darson FineFarhati MardhiyahNo ratings yet
- Metabolism MCQ ADocument0 pagesMetabolism MCQ ASurajit BhattacharjeeNo ratings yet
- Fatty Acid Synthesis 11.12.19Document18 pagesFatty Acid Synthesis 11.12.19Sanreet Randhawa100% (1)
- Carbohydrate MetabolismDocument30 pagesCarbohydrate MetabolismAndraKamee100% (1)
- CHAPTER 18 GlycolysisDocument10 pagesCHAPTER 18 Glycolysis楊畯凱No ratings yet
- Glycolysis NotesDocument2 pagesGlycolysis NotesAnsley Ellen BarfieldNo ratings yet
- Cardiac MetabolismDocument35 pagesCardiac Metabolismwaqas_xsNo ratings yet
- GlycogenolysisDocument37 pagesGlycogenolysisJasmine Rey QuintoNo ratings yet
- Metabolism of DisaccharidesDocument15 pagesMetabolism of DisaccharidesminaNo ratings yet
- Glycogenesis: DR Nazakat HussainDocument16 pagesGlycogenesis: DR Nazakat HussainMuhammad Jamal100% (1)
- Metabolism of Purine & Pyrimidine NucleotidesDocument38 pagesMetabolism of Purine & Pyrimidine NucleotidesShimmering MoonNo ratings yet
- بايو بيبر 1 د.جميلة السمهريDocument8 pagesبايو بيبر 1 د.جميلة السمهريOzgan SüleymanNo ratings yet
- Metabolism of Carbohydrate: Department of Biochemistry Faculty of Medicine University of YARSI JakartaDocument60 pagesMetabolism of Carbohydrate: Department of Biochemistry Faculty of Medicine University of YARSI JakartaAmanda PutriNo ratings yet
- Fa Lipid MetabDocument4 pagesFa Lipid MetabJoy VergaraNo ratings yet
- Digestion and Absorption of Carbohydrates: by Arpitha 3 BnysDocument31 pagesDigestion and Absorption of Carbohydrates: by Arpitha 3 BnysBig ZeroNo ratings yet
- Carbohydrate Metabolism SamplexDocument13 pagesCarbohydrate Metabolism SamplexCherie ThompsonNo ratings yet
- Clinical SignificanceDocument31 pagesClinical Significancejav israelNo ratings yet
- Nucleotide Biosynthesis Lecture 2017sDocument23 pagesNucleotide Biosynthesis Lecture 2017sSaad KazmiNo ratings yet
- Git Biochemistry of GitDocument9 pagesGit Biochemistry of GitSibatNo ratings yet
- Carbohydrates MetabolismDocument6 pagesCarbohydrates MetabolismAisha ShahfiqueeNo ratings yet
- Citric Acid Cycle - Pyruvate DehydrogenaseDocument33 pagesCitric Acid Cycle - Pyruvate Dehydrogenasesultan khabeeb100% (1)
- Metabolism of CarbohydrateDocument48 pagesMetabolism of CarbohydrateAbdullah TheNo ratings yet
- Integration of Metabolism Integration of MetabolismDocument10 pagesIntegration of Metabolism Integration of MetabolismEdison LucianoNo ratings yet
- (مكتملة) اسئلة البايو د.ج السمهري (.)Document10 pages(مكتملة) اسئلة البايو د.ج السمهري (.)Ozgan SüleymanNo ratings yet
- MCQ MembranesDocument10 pagesMCQ MembranesMarilyne RizkNo ratings yet
- 1st Long Quiz - Overview of Metab, Carb Metab 1 and 2Document5 pages1st Long Quiz - Overview of Metab, Carb Metab 1 and 2MineTagraNo ratings yet
- Integration of Metabolism 2019Document68 pagesIntegration of Metabolism 2019Cahyani Tiara Safitri100% (1)
- Glycolysis PresentationDocument34 pagesGlycolysis PresentationAl LimNo ratings yet
- SummaryDocument21 pagesSummarydindaNo ratings yet
- Cholesterol - Synthesis, Metabolism, Regulation PDFDocument10 pagesCholesterol - Synthesis, Metabolism, Regulation PDFAdreiTheTripleA100% (1)
- Hexose Monophosphate ShuntDocument43 pagesHexose Monophosphate ShuntSecret AgentNo ratings yet
- Glycolysis ProcessDocument7 pagesGlycolysis ProcessBlister CountNo ratings yet
- Biochemistry VivaDocument8 pagesBiochemistry VivaNazia WasimNo ratings yet
- Bibc102, Metabolic Biochemistry: Gen-Sheng Feng Fall, 2010Document14 pagesBibc102, Metabolic Biochemistry: Gen-Sheng Feng Fall, 2010cool_trainerNo ratings yet
- Glycoproteins and Proteoglycans TTVDocument34 pagesGlycoproteins and Proteoglycans TTVcraigNo ratings yet
- LO9&10Document13 pagesLO9&10Nine AranasNo ratings yet
- 9 Bacterial MetabolismDocument7 pages9 Bacterial MetabolismAlphonse Rossaint SambranoNo ratings yet
- 8787 37898 1 PBDocument4 pages8787 37898 1 PBGabriella NaomiNo ratings yet
- Photosynthesis PDFDocument20 pagesPhotosynthesis PDFShammahNo ratings yet
- C 1 1 2025 Topic Test MsDocument4 pagesC 1 1 2025 Topic Test MsRawanMazen SharifNo ratings yet
- Ingles 2 Carbohidratos PDFDocument4 pagesIngles 2 Carbohidratos PDFIsabela OrihuelaNo ratings yet
- '-Daftar Harga Reagen LabtestDocument1 page'-Daftar Harga Reagen LabtestUPT Labkes R/LNo ratings yet
- The Hill RXN in Isolated Chloroplasts PostDocument12 pagesThe Hill RXN in Isolated Chloroplasts PostKarina Khan100% (12)
- Chapter 10 Optional HomeworkDocument19 pagesChapter 10 Optional Homeworkcolahmundu100% (7)
- Carbohydrate Metabolism-1Document27 pagesCarbohydrate Metabolism-1RESHMA I MBANo ratings yet
- Biochemistry 4th Edition Mathews Test Bank 1Document36 pagesBiochemistry 4th Edition Mathews Test Bank 1loribowersbwknifoyxe100% (23)
- Diabetes Mellitus MCQsDocument4 pagesDiabetes Mellitus MCQsvaidehianand007No ratings yet
- Plickers Example Data Per ClassDocument2 pagesPlickers Example Data Per Classapi-377689746No ratings yet
- Biology Lab #12 Discussion (HOW BREATHING RATE CHANGES WITH EXERCISE)Document3 pagesBiology Lab #12 Discussion (HOW BREATHING RATE CHANGES WITH EXERCISE)Joy-Ann NuptialNo ratings yet
- Quarter 1 Week 7 Science 9Document38 pagesQuarter 1 Week 7 Science 9Denmiel MagnayeNo ratings yet
- Cellular Respiration-BioDocument35 pagesCellular Respiration-BioKarylle Chloe MarceloNo ratings yet
- Acetyl Coenzyme A - AssignmentDocument2 pagesAcetyl Coenzyme A - AssignmentSasa LiliNo ratings yet
- Chapter 6 Active Reading GuideDocument6 pagesChapter 6 Active Reading GuideSoofia KhanNo ratings yet
- Glycolysis: Cellular Respiration Cells CytoplasmDocument4 pagesGlycolysis: Cellular Respiration Cells CytoplasmArav PatelNo ratings yet
- Las Q4 Week-2 GenDocument8 pagesLas Q4 Week-2 GenFranzhean Balais CuachonNo ratings yet
- Bob's - Children's Book ProjectDocument11 pagesBob's - Children's Book ProjectRunningLightxNo ratings yet
- Photosynthesis: Study Guide Photosynthesis Equation: 6CO2 + 6H2O - C6H12O6 + 6O2Document4 pagesPhotosynthesis: Study Guide Photosynthesis Equation: 6CO2 + 6H2O - C6H12O6 + 6O2vscolegit shoppeNo ratings yet
- GlycolysisDocument21 pagesGlycolysisAli AlqumaNo ratings yet
- Biology S6Document10 pagesBiology S6AKAYEZU Body santiveNo ratings yet
- Chapter Ix - Mechanism of Lipids MetabolismDocument33 pagesChapter Ix - Mechanism of Lipids MetabolismAngelo AngelesNo ratings yet
- 2.glycolysis & Oxidation of PyruvateDocument12 pages2.glycolysis & Oxidation of Pyruvateقتيبه خالد دحام خلفNo ratings yet
- G9 C4-1 - Plant PhotosynthesisDocument14 pagesG9 C4-1 - Plant Photosynthesiskatherine andinoNo ratings yet
- LMSG 10Document6 pagesLMSG 10Jasmen Mohawk CheaseNo ratings yet
- BIO1 Lesson-12 PhotosynthesisDocument36 pagesBIO1 Lesson-12 PhotosynthesisCath Detoperez100% (1)
- Análise Comparativa Do Gasto Energético Entre As Equações de Harris-Benedict e de Long e A Calorimetria Indireta em Pacientes SépticosDocument8 pagesAnálise Comparativa Do Gasto Energético Entre As Equações de Harris-Benedict e de Long e A Calorimetria Indireta em Pacientes SépticosJoão Lucas SimõesNo ratings yet
- Photosynthesis - Part 1Document90 pagesPhotosynthesis - Part 1Antonio WilloughbyNo ratings yet