You might also like
- Propuesta para Una Buena AlimentacionDocument21 pagesPropuesta para Una Buena AlimentacionDavidHidalgoPonceNo ratings yet
- Disfagia OrofaríngeaDocument23 pagesDisfagia OrofaríngeaClau Guzmán MartínezNo ratings yet
- Diseccion de RanaDocument5 pagesDiseccion de Ranaarle_mendozaNo ratings yet
- Resumen Trastorno de La PersonalidadDocument4 pagesResumen Trastorno de La PersonalidadPPNo ratings yet
- Manual General HISDocument18 pagesManual General HISkeymaker008767% (6)
- Cuando Los Adolescentes y Los Adultos Jóvenes EstánDocument3 pagesCuando Los Adolescentes y Los Adultos Jóvenes EstánCM RunneeyNo ratings yet
- Penetracion Digestion Energias PerversasDocument21 pagesPenetracion Digestion Energias PerversascarloscappNo ratings yet
- Papillomavirus-Associated Cutaneous Neoplasia in MDocument13 pagesPapillomavirus-Associated Cutaneous Neoplasia in MValentina Turra GallardoNo ratings yet
- Examen Radiografico Dental Medicina IIDocument27 pagesExamen Radiografico Dental Medicina IIlcomar_ftoooooNo ratings yet
- Oftalmologia Pediatrica para Todos Los DiasDocument14 pagesOftalmologia Pediatrica para Todos Los DiasRoberto FajardoNo ratings yet
- Cerebro y EmociónDocument36 pagesCerebro y EmociónWillir100% (1)
- Bacteriologia PracticaDocument7 pagesBacteriologia PracticaFernando Silva MichalonNo ratings yet
- Caso N ° 2 Taller PDFDocument19 pagesCaso N ° 2 Taller PDFGatica NataliaNo ratings yet
- Matriz de RiesgosDocument32 pagesMatriz de RiesgosMauricio YaselgaNo ratings yet
- Cáncer de PróstataDocument6 pagesCáncer de PróstataholaNo ratings yet
- Clase 4 Sinapsis Act.-1Document39 pagesClase 4 Sinapsis Act.-1Roel VaNo ratings yet
- La Psicopatia Diferencia Con TPASDocument24 pagesLa Psicopatia Diferencia Con TPASRoberto GalleguillosNo ratings yet
- Protocolo Hipertension Pulmonar 2019 PDFDocument144 pagesProtocolo Hipertension Pulmonar 2019 PDFManes Denis100% (1)
- Mesoterapia Facial 2Document7 pagesMesoterapia Facial 2Luis S0% (1)
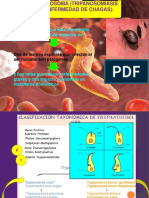
- ChagasDocument66 pagesChagasluceritoNo ratings yet
- Teorias de EnfermeriaDocument14 pagesTeorias de EnfermeriaJesús FuentesNo ratings yet
- SangreDocument3 pagesSangrelauraNo ratings yet
- Páginas desdeLIBRO PDFDocument2 pagesPáginas desdeLIBRO PDFmdmunozc01No ratings yet
- Examen Oral de Biologia 1º MedioDocument17 pagesExamen Oral de Biologia 1º MedioCarolina Torres ArmijoNo ratings yet
- RP-CTA1-K01 - Ficha #7Document7 pagesRP-CTA1-K01 - Ficha #7EroicoMosquera100% (1)
- Enfermedad de Graves Basedow IntratableDocument1 pageEnfermedad de Graves Basedow IntratableKtrym BustosNo ratings yet
- Métodos de Diagnostico en ParasitologiaDocument2 pagesMétodos de Diagnostico en Parasitologialuiqui7No ratings yet
- Atención de Enfermería en Admisión HospitalariaDocument3 pagesAtención de Enfermería en Admisión HospitalariaAdarely CP43% (7)
- Alteraciones de La QueratinizacionDocument20 pagesAlteraciones de La Queratinizacionadriana barajas0% (1)